XiaoMi-AI文件搜索系统
World File Search System神经元

在体外神经元发育形态的半自动定量评估
成熟的神经元表现出其轴突和树突(统称为神经突)的广泛树皮化,以与相邻细胞形成功能连接并接收感觉信号。独特的神经元结构被认为会引起神经元的计算能力(Cuntz,Borst,&Segev,2007; Ferrante,Migliore和Ascoli,&Ascoli,2013; Kanari等人。,2018年; Van Elburg&van Ooyen,2010年; Zomorrodi,Ferecsk´o,Kov´acs,Kréoger和Timofeev,2010年)。In addition, morphological differences between neuronal cell types are thought to result in their functional differences ( Khalil, Farhat, & Dl otko , 2021 ; Krichmar, Nasuto, Scorcioni, Washington, & Ascoli , 2002 ; Mainen & Sejnowski , 1996 ; Schaefer, Larkum, Sakmann, & Roth , 2003 ; Vetter, Roth, & h ausser,2001年)。在体外原发性神经元中这种关键结构的发展过程中,几种形态学变化已被归类为不同的阶段,这些阶段可以定性地描述(Dotti,Sullivan,&Banker,1988; Powell,Rivas,Rodriguez-Boulan,&Hatten,&Hatten,&Hatten,1997; Tahirovic&Bradke&Bradke,2009年)。

MDM的Ivanti神经元96管理员指南
About Ivanti Neurons for MDM 5 New features summary 6 General features and enhancements 6 Android features 7 iOS, macOS, and tvOS features 8 Windows features 11 Mobile Threat Defense features 11 Getting Started 12 Solution Overview 12 Setting preferred language in a browser 18 Unified navigation interface for Ivanti Neurons for MDM and Access 19 Device Admin (DA) mode to manage Android devices - deprecated 19 Configuring macOS devices 21 Configuring and using registration confirmation emails 26 Configuring and using policy compliance notification emails 27 On-demand features 29 Preparing for Android Enterprise device support 33 Dashboard 36 Working with Widgets 37 App Insights 52 Using Scheduled Reports 58 Using Custom Reports 69 Users 80 Adding Users 81 User Groups 87 User Settings 91 User Branding 107 User Enrollment with Apple Business Manager 109 Account driven User Enrollment 121 User Licenses 123 Managing用户124个设备168从设备开始使用169个设备组188非管理设备195 App库存197管理设备201 Apps 288

Notch信号在调节神经元中起双重作用
神经祖细胞会产生兴奋性神经元,其次是少突胶质细胞(OLS)和垂体细胞。然而,调节该神经元时间 - 胶质开关的特定机制尚未完全了解。在这项研究中,我们表明,在胚胎发育的后期阶段,需要在背前祖细胞中Notch信号的适当平衡才能产生少突胶质细胞。在两性的小鼠胚胎中使用离体和子宫方法中,我们发现Notch抑制减少了背胸膜中少突胶质细胞的数量。然而,缺口过度活化也阻止了寡构成并保持祖细胞状态。这些结果表明,在促进和抑制寡头生成中,Notch信号传导的双重作用,必须对其进行微调才能在正确的时间和正确的数字中生成少突胶质细胞谱系细胞。在此过程中,我们进一步将其下游的典型档位hes1和hes5确定为负调节剂。crispr(群集定期间隔短的短质体重复)/cas9介导的hes1和hes5的敲低敲低导致促寡胶质细胞因子ASCL1的表达增加,并导致早产性寡构成。相反,将缺口与ASCL1过表达结合起来,可稳健地促进寡头生成,表明与ASCL1合成的Notch机制单独的机制,以指定少突胶质细胞的命运。我们提出了一个模型,其中Notch信号与ASCL1一起工作以指定祖细胞朝向少突胶质细胞谱系,但也通过hES依赖ASCL1的抑制来维持祖细胞状态,从而使少突胶质细胞不太早,从而导致神经元的精确时间促成神经元 - Glia Switch。

终身患运动神经元疾病的风险(...
终生发展运动神经元疾病(MND)的风险约为300中的1个。该疾病的确切原因仍然未知,并且需要进行研究的资金来理解这一点,并最终需要开发治疗。MND协会致力于照顾与MND的人,并通过我们的研究资金来照顾和影响MND的世界。先前的研究试图确定运动或对接触运动的运动影响是否导致神经系统疾病的风险增加,例如阿尔茨海默氏病,MND和慢性创伤性脑病(CTE)。在过去的十年中,一些证据表明,包括足球和美式足球(NFL)的接触运动之间存在潜在的联系,而神经退行性疾病的风险增加了。的发现表明,在专业水平上踢足球的人患神经退行性疾病的风险增加,其中一些研究表明,成为职业足球运动员会增加一个人发展MND的风险(Chio等2005:Chio等人,Chio等,2009)。另一项研究(Russell等,2022年)得出结论,与普通人群相比,前苏格兰国际橄榄球联盟球员患MND的风险增加了。也有少量的研究发现,表明对MND的敏感性增加与某些专业运动有关,并且运动容易重复脑震荡头和颈脊柱创伤(Blecher等人,2019年)。当然,这项研究在MND协会中引起了人们的极大兴趣,公众对接触运动和神经退行性疾病的风险的关注正在增加。尽管这些研究迄今进行了这些研究表明这些职业运动与MND之间的相关性并未证明因果关系 - 因此他们认识到,职业足球运动员和橄榄球运动员更有可能发展MND,但他们并不建议这样做专业地踢足球,或者直接导致一个人发展MND。也要牢记这些研究中报告的MND病例的数量仍然相对较低,因此得出结论,如果由于随机的机会,这只是一个群集,则可能会误解风险的确定风险。尽管体育内部的运动越来越多,可以理解,认识并在必要时减轻负面的长期影响,但也很明显,需要更多的研究。我们认识到这一点,并正在与研究人员和机构合作,以促进MND研究。我们已经与苏格兰MND慈善机构和我的名字“ Doddie基金会”合作,以资助新的研究,以研究创伤性脑损伤是否会导致发展MND的风险增加。这项研究,创伤性脑损伤和运动神经元疾病(T-MND)由威廉·斯图尔特教授及其团队在格拉斯哥大学领导。我们也是MND和Sport Expert Working Group的一部分,该组织致力于探索Sport和MND之间的潜在联系。该小组包括MND协会,MND Scotland和我的名字“ Doddie以及MND的人们)的代表,并且正在回应许多科学出版物,这些出版物表明,在

对皮质投影神经元命运多样性的时间控制
摘要在新皮层发育过程中,皮质投射神经元(PN)被顺序产生并组装成我们与环境相互作用的基础的电路。皮质PN在出生日期,层位置,近距离身份,连通性和功能方面是异质的。这种多样性在进化最新的物种中引起了争议,但是在皮质生成期间何时以及如何出现。虽然确定基因和早期随机性的时间锁定表达允许产生不同类型的PN,但在展开相似的转录过程中的时间差异,而不是这些程序中的基本差异,但进一步说明了PN亚型和跨物种之间的解剖变异性。总的来说,这些机制将在此处讨论,它参与了增加皮质PN多样性。
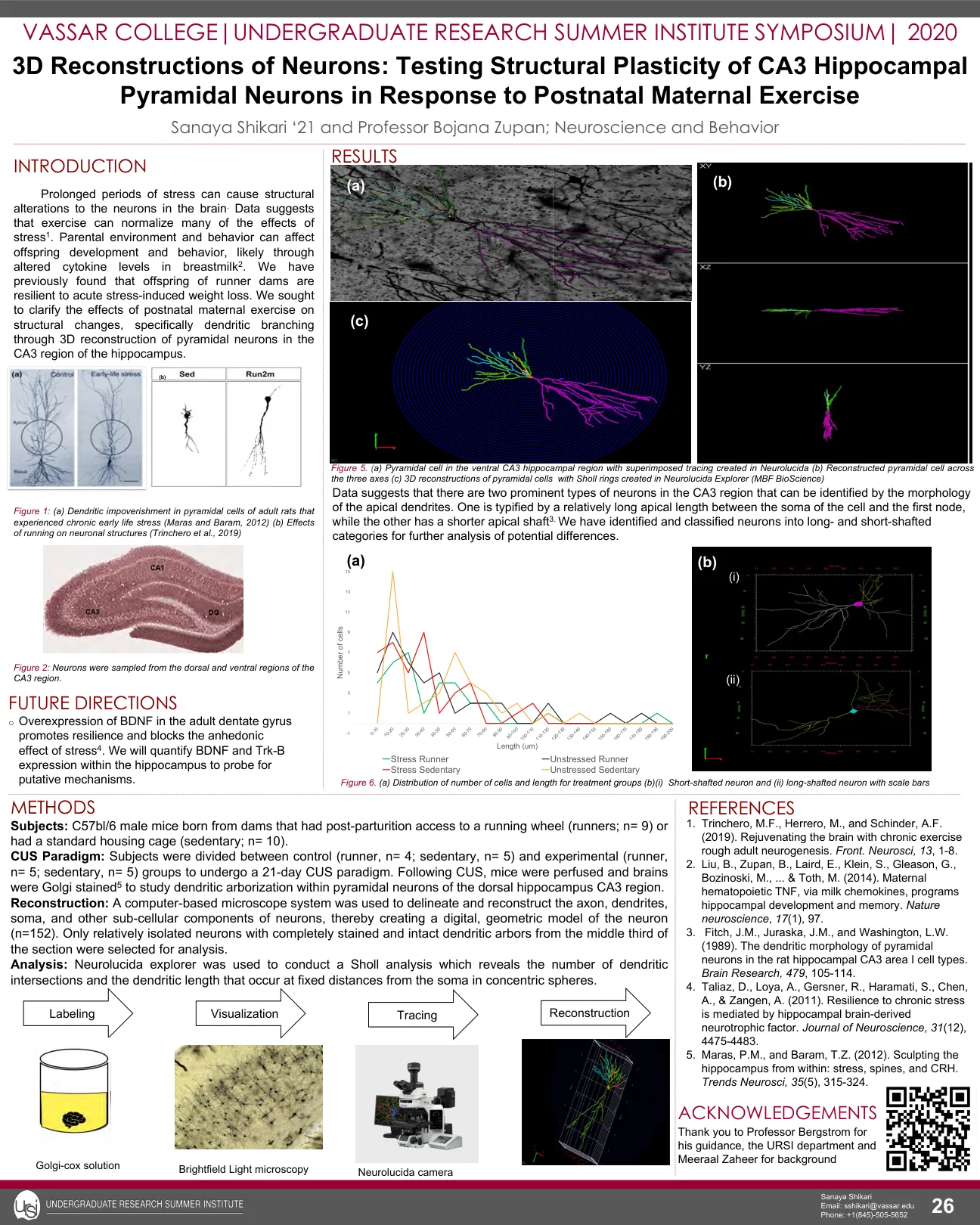
神经元的 3D 重建:测试 CA3 海马锥体神经元对产后母亲运动的反应的结构可塑性
方法 受试者:C57bl/6雄性小鼠,其母鼠产后可使用跑轮(跑步者;n= 9)或使用标准笼子(久坐;n= 10)。 CUS 范式:将受试者分为对照组(跑步者,n= 4;久坐组,n= 5)和实验组(跑步者,n= 5;久坐组,n= 5),接受为期 21 天的 CUS 范式。 CUS 之后,对小鼠进行灌注,并对大脑进行 Golgi 染色 5,以研究背海马 CA3 区锥体神经元内的树突树枝状化。 重建:使用基于计算机的显微镜系统来描绘和重建神经元的轴突、树突、胞体和其他亚细胞成分,从而创建神经元的数字几何模型(n=152)。仅选择切片中间三分之一处具有完全染色和完整树突状体的相对分离的神经元进行分析。分析:使用 Neurolucida explorer 进行 Sholl 分析,该分析揭示了同心球中距胞体固定距离处出现的树突交叉点数量和树突长度。

神经形态感知的有机电化学神经元
摘要尽管在工作生活中广泛引入数字技术以及对包括学校在内的公共部门的需求,但数字化与工作环境之间的关系的研究有限。因此,本文的目的是探讨数字行政和通信系统和教学工具的实施过程如何与学校经历的与工作相关的需求和资源互动。使用标准化过程理论(NPT)和工作需求资源模型(JDR)来分析瑞典学校的数字化经验以及员工,经理,战略家以及健康与安全官员的工作环境。根据对25名参与者的半结构化访谈,这项研究表明,在引入数字技术时,资源缺乏和太高的工作量是有问题和挑战性的。合作合作和决策机会似乎增强了数字化过程,甚至在某种程度上甚至在某种程度上桥梁就弥合了紧张的时间资源和高工作量的弱点。

脊椎动物全局神经元工作区的进化起源
意识的全局神经元工作空间理论提供了一种明确的功能架构,将意识与认知能力(如感知、注意力、记忆和评估)联系起来。我们表明,主要基于人类研究的全局神经元工作空间的功能架构与描述最小意识的无限联想学习理论提出的认知情感架构相对应。然而,我们认为,当应用于基础脊椎动物时,这两个模型都需要进行重要的修改,以适应人们对脊椎动物大脑进化的了解。最重要的是,比较研究表明,在基础脊椎动物中,全局神经元工作空间是由海马同源物中的事件记忆系统实例化的。这一提议对于理解海马和皮质功能、记忆与意识的进化关系以及统一感知的进化具有可检验的预测和意义。

发现神经元中的新型转录增强子...
神经精神疾病越来越普遍。鉴于其复杂且多因素的发病机理,迫切需要有效且有针对性的疗法可以改善患者的生活质量。全基因组关联研究(GWASS)已经确定了各种遗传改变,这些改变有助于神经精神疾病的发展和发展,从轻度阅读障碍到更严重的疾病,例如精神分裂症。虽然成千上万的单核苷酸多态性(SNP)(SNP)与DNA中的单个核苷酸位置发生了变化 - 与神经系统疾病有关,但大多数位于基因组的非编码区域。尽管这些非编码区未编码蛋白质,但它们包含调节元素,例如增强子序列,在控制基因表达中起着至关重要的作用。增强子可以在长距离内调节基因活性,并且通常特定于细胞类型和发育阶段。尽管其重要性,但增强子的特征仍然很差,并且尚未完全了解其在神经系统发展和疾病中的精确功能。在一项新的研究中,奇巴大学高级学术研究与医学研究院医学研究所Masahito教授以及Karolinska Institutet,Sweden,Sweden和PelinSahlénnewlobleInstutter from fromniwleart Institute froment from Technology的Karolinska Institutet的Huddinge(MedH)的Juha Kere和Peter Swoboda教授以及彼得罗斯卡研究所(Karolinska Institutet)的彼得·斯沃博达(Peter Swoboda)博士。他们还研究了与神经元疾病有关的假定增强子与GWAS识别的基因座之间的关联。他们进行了一系列高级分析,以使用Luhmes细胞来识别和表征参与神经元分化的增强子,Luhmes细胞是源自人类胎儿中脑多巴胺能神经元的细胞系。该研究的主要作者Yoshihara博士很快就会发表在EMBO报告中,他说:“阐明与疾病相关的变体影响基因调节的方式可以揭示以前统一的参与神经元疾病的分子途径,并揭示了用于药物开发的新型治疗靶标。”研究人员使用了luhmes神经元前体细胞,这些细胞可以分化为与人脑衍生神经元具有高转录相似性的功能性神经元。他们采用了基因表达(CAGE)和天然伸长转录本(净)键的CAP分析,以识别和量化基因组宽水水平的启动子和增强子的活性。这些技术与靶向的染色体构象捕获(Capture Hi-C/HICAP)相结合,这是一种将远处增强子与其靶基因联系起来的高级测序方法。该分析确定了47,350个主动推定增强剂,其中65.6%是新颖的,并且证明了与帕金森氏病,精神分裂症,双相情感障碍和主要抑郁症相关的SNP富集。最后,他们在培养细胞中进行了体外测定,以验证启动子增强子相互作用。使用CRISPR-CAS9系统进行基因组编辑,他们激活了与神经元分化和疾病有关的基因的增强子和启动子。与他们的分析一致,增强子的激活导致靶基因的表达水平显着升高。

损伤后神经元再生:基因治疗的新视角
脊髓损伤 (SCI) 是全球范围内导致残疾的主要原因,再生医学为开发此类损伤的新疗法带来了希望 ( James et al., 2019 )。SCI 可导致感觉和运动功能丧失,并可能对个人的生活质量产生重大影响,不仅影响身体能力,还影响情绪和社会健康 ( Eckert and Martin, 2017 )。尽管经过数十年的研究,但 SCI 仍然无法治愈。脊髓受损神经元无法再生是再生医学领域的主要挑战之一。在哺乳动物中,脊髓是一种复杂的结构,再生能力有限 ( He and Jin, 2016 ; Sofroniew, 2018 ),调节神经元再生的细胞和分子机制尚不完全清楚。最近的研究确定了促进神经元再生的新靶点和潜在策略,包括使用干细胞疗法(Okano,2010 年;Führmann 等人,2017 年)、基因疗法(Lentini 等人,2021 年;Zhang Y. 等人,2022 年)和组织工程(Madhusudanan 等人,2020 年;Cheng 等人,2021 年)。最近的研究强调了使用基因疗法促进各种情况下的再生和功能恢复。例如,通过免疫逃逸强力霉素诱导基因开关使用时间限制的神经胶质细胞系衍生的神经营养因子表达的基因疗法已显示出在增强大鼠近端神经损伤后的轴突再生和运动神经元存活方面的前景(Eggers 等人,2019 年)。研究表明,在 SOX2 介导的体内命运重编程后,驻留的星形胶质细胞会生成新的神经元(Su 等,2014;Wang 等,2016)。同样,另一项研究表明,NG2 神经胶质细胞中的异位 SOX2 可诱导神经发生、减少神经胶质瘢痕形成并生成脊髓本体神经元,促进功能恢复(Tai 等,2021)。此外,研究表明,脊髓损伤后进行 FGF22 基因治疗可促进突触形成并为神经元重新布线提供有针对性的支持,急性和早期应用可改善功能恢复(Aljovi´c 等,2023)。然而,结果显示存在一个较短的时间范围,至少在 SCI 后的最初 24 小时内,在此期间,使用 FGF22 进行突触形成基因治疗可以改善运动功能的恢复。这种有限的窗口在临床环境中可能难以实现,这可能需要探索具有更长治疗窗口的替代突触生成分子或方法。总体而言,这些发现表明基因疗法有可能激活内源性神经胶质细胞的再生能力,从而导致各种情况下的再生和功能恢复。