机构名称:
¥ 1.0
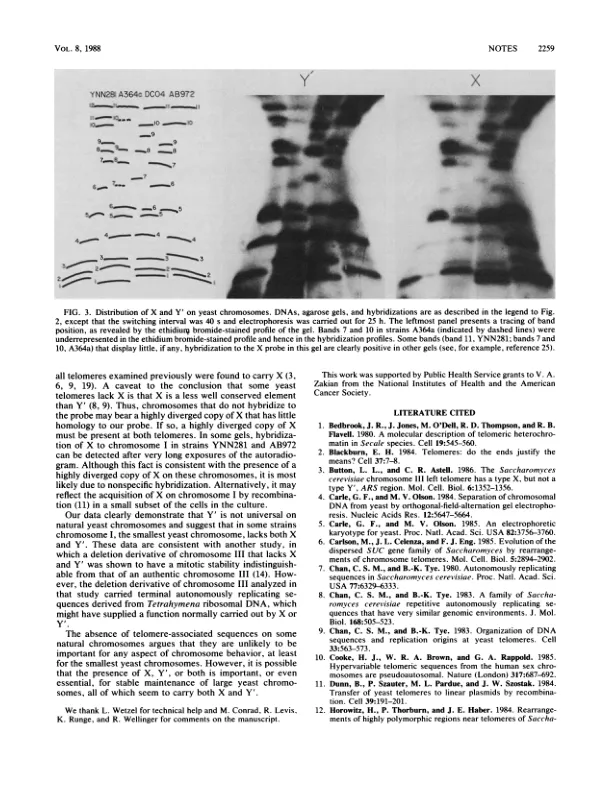
关于端粒区的结构,一个共同的主题正在出现。染色体末端带有多个串联重复的简单卫星状 DNA(2)。除了染色体末端的简单序列外,端粒附近的区域通常还带有长段中间重复 DNA(1、10、13、15、18、24)。在酿酒酵母中,染色体以 200 到 600 个碱基对的不规则序列 C1_3A 结束(17、23;图 1)。此外,在 DNA 末端附近发现了两个中间重复元素,称为 X 和 Y'(8、9)。Y' 是一个高度保守的元素,长度为 6.7 千碱基(kb)(8、9)。 X 是一种比 Y' 保守性更低的元件,大小范围为 0.3 至 3.75 kb,位于 Y' 的着丝粒附近(8, 9)。C1_3A 重复序列的内部序列以及 DNA 复制的推定起点(自主复制序列)与 X 和 Y' 相关(7, 21)。这些特性与端粒相关序列在复制、重组或端粒区域修复中发挥作用相一致。已经开发出凝胶系统,可以分离完整的酵母染色体 DNA 分子(4, 16)。已记录了菌株 YNN281、A364a、DCO4 和 AB972(5)中每条染色体在一个系统(正交场交替凝胶电泳 [OFAGE])中的行为。通过改良的凝胶插入法 (16) (5) 从这些菌株中制备 DNA,并进行 OFAGE 处理。将 DNA 转移到硝酸纤维素上并与 X 和 Y' 特异性探针杂交 (20)(图 2)。通过琼脂糖凝胶分离 1.7 kb NcoI 片段,从 YRp12O (9) 制备 X 特异性探针。通过分离 1.7 kb BglII 片段,从 YRpl31b (9) 制备 Y' 特异性探针,该片段被亚克隆到 BamHI 消化的 M13 mpl8 中。从 pYtl03 (17) 切下 125 碱基对 HaeIII-MnlI 片段,其中包含 82 碱基对 C1_3A 重复序列。杂交探针来自据报道不含 C1_3A 重复序列的 X 和 Y' 区域。这一点已通过以下事实得到证实:源自 pYtl03 的真正的 C1_3A DNA 既不与 X 也不与 Y' 探针杂交。为探针选择的 X 区域在不同的 X 元素中是保守的 (8, 9)。表 1 中显示的数据是从 17 种不同的凝胶中汇编而来的,这些凝胶的切换间隔范围为 20 到 80 秒。每个菌株的 X 和 Y' 分布模式不同(图 2 和 3)。每个菌株中至少有三条最小染色体中有一条不与 Y' 探针杂交,在三个菌株中,五条最小染色体中的两条不与 Y' 探针杂交
自然界中端粒相关序列的分布
主要关键词



相关文件推荐