XiaoMi-AI文件搜索系统
World File Search SystemC57BL

TDG CAS9-CKO策略
过程如下:在体外转录GRNA。cas9和grnas微注射到C57BL/6JGPT小鼠的受精卵中。受精卵被移植以获得通过PCR和靶标扩增子测序确认的阳性F0小鼠。通过与C57BL/6JGPT小鼠配对阳性F0产生小鼠获得稳定的F1生成小鼠菌株,并通过PCR和靶向Agplicon测序对所需的突变等位基因进行确认。

材料的最新进展是否存在磁性单孔吗?
应用于产生基因组编辑的大鼠,包括白化病sprague-dawley和白化病刘易斯大鼠(但是,不是有色的棕色挪威[bn]大鼠)。我们观察到成功的I -Gonad取决于所使用的小鼠菌株。例如,在随机繁殖小鼠(例如ICR和C3H/HE×C57BL/6)中,它在相对严格的电气条件下成功,但在C57BL/6菌株中却没有成功。在不太严格的条件下,I -Gonad在C57BL/6菌株中取得了成功。我们推测使用BN大鼠对I -Gonad也是如此。在应用> 500 mA的电流时,我们未能获得大鼠后代(胎儿/新生儿);但是,使用NEPA21(NEPA基因)在100-300 Ma下I-Gonad导致基因组编辑的BN大鼠的产生,其效率为75%-100%。同样,使用CUY21EDIT II(BEX Co.)在150-200 Ma的电流下,I-Gonad导致基因组编辑的BN大鼠的产生,其效率为24%-55%。这些实验表明,在执行I -Gonad时,根据所使用的大鼠菌株选择适当的电流值的重要性。

利用专有脂质实现体内基因编辑...
• 小鼠品系:C57BL/6 和 ApoE KO • 剂量:1 mg/kg • Life Edit LNP • mRNA:fLuc + b-gal 组合 (1:1) • 时间点:静脉注射后 6 小时

使用 Split AAV 进行体内遗传性眼病矫正-...
体外:293T、人类视网膜类器官、iPSC 衍生的 RPE、人类视网膜外植体、人类 RPE/脉络膜外植体 体内:C57BL/6J 小鼠(WT 和 Abca4 huG1961E)、非人类灵长类动物(食蟹猴)

大脑输送平台的合理设计 - BioArctic
正常小鼠水平(C57BL/6):红细胞:9.32 0.7(10 12 /L)血红蛋白; 14.7 1.08(g/dL)血细胞比容:44.0 3.5(%)(Talbot 等,PLoS ONE 6(8) (2011))

引用:McGrath KC、Li X、Twigg SM、Heather AK (2020) 载脂蛋白-AI 模拟肽 D-4F 和 L-5F 可减轻 C57BL/6 小鼠的肝脏炎症并增加胰岛素敏感性。PLoS ONE 15(1): e0226931。 https://doi.org/10.1371/ journal.pone.0226931
高密度脂蛋白 (HDL) 的抗炎和抗氧化特性的发现引发了一个问题:HDL 是否可用于治疗疾病中的炎症。胰岛素抵抗主要依赖于肝脏炎症,研究表明,组成性活性肝核因子 κB (NF- κ B)(驱动炎症反应的中枢介质)会导致小鼠模型中的胰岛素抵抗状态 [ 1 ]。我们之前证明,单独(无脂质)或作为含有 PLPC 的重组 HDL(rHDL)的一部分施用载脂蛋白 A-I (apoA-I) 可预防肝脏炎症并改善高脂喂养的 C57Bl/6 小鼠的胰岛素抵抗 [ 2 ]。虽然结果令人鼓舞,但 apoA-I 颗粒的产生很大且耗时。载脂蛋白 A-I (apoA-I) 模拟肽已被开发和设计,其功能类似于全长 apoA-I,但效力更强,药代动力学特征也比全长 apoA-I 更好 [ 3 , 4 ]。这些模拟肽比 rHDL 有许多优势,包括成本相对较低、生产简单,并且能够修改其结构以允许口服给药。ApoA-I 模拟肽的特点是其苯丙氨酸残基附着,苯丙氨酸残基的增加与疏水性的增加及其与磷脂结合的能力相关 [ 4 ]。根据序列中疏水性苯丙氨酸残基的数量,研究最深入的是 4F 和 5F apoA-I 模拟肽。它们具有与 apoA-I 相同的 A 类两亲性螺旋结构 [ 5 ]。D-4F 和 L-5F 的区别在于两亲性螺旋疏水面上存在的苯丙氨酸数量,并且 D-4F 由 D-氨基酸合成,而 L-5F 由 L-氨基酸合成。对于口服给药,与 L-氨基酸相比,D-氨基酸被发现更能抵抗酶促降解。然而,吸收后,D-氨基酸不会降解,这可能导致毒性和其他副作用 [ 6 ]。L-氨基酸在循环中的毒性较小,但它们不能抵抗口服的酶促降解。两者都曾在生物学研究中使用过,据报道具有强大的抗炎和抗氧化作用 [ 7 – 12 ]。我们表明,模拟治疗显着在本研究中,我们直接测试了使用 apoAI 模拟肽 D-4F 和 L-5F 治疗是否可以改善高脂饮食 (HFD) 喂养小鼠的胰岛素敏感性,从而降低肝脏炎症。
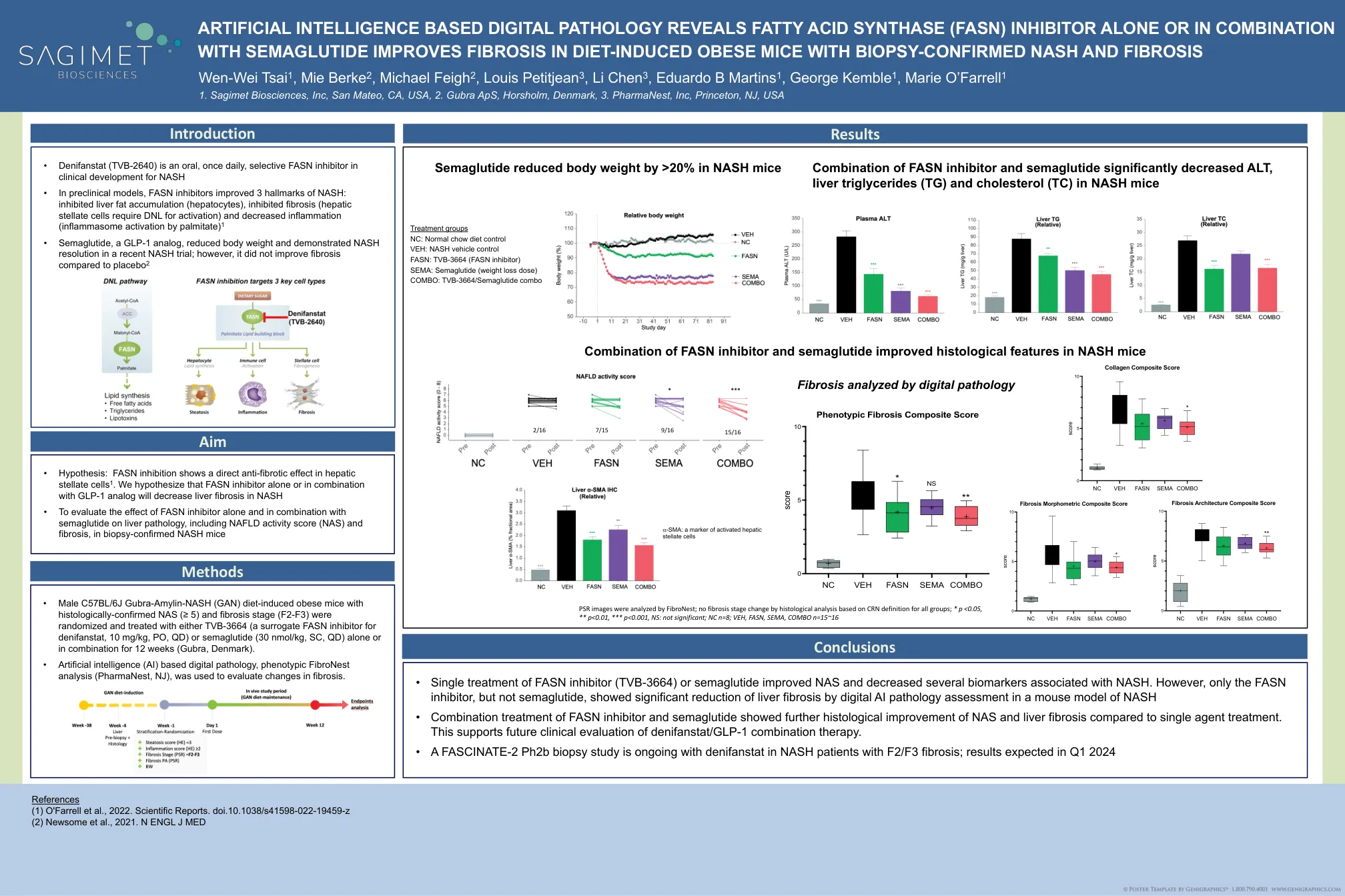
基于人工智能的数字病理学揭示了脂肪...
•雄性C57BL/6J Gubra-氨基蛋白-NASH(GAN)饮食诱导的肥胖小鼠,具有组织学确认的NAS(≥5)(≥5)和纤维化阶段(F2-F3)(F2-F3),并用TVB-3664进行治疗,并用TVB-3664治疗(替代fasn infasn i抑制剂,nefanstattate demifate in if nifanStat,30 mg/kg,或30 mg/kg,或quut),或者NMOL/KG,SC,QD)单独或组合12周(丹麦Gubra)。

BI特异性自身抗原T细胞诱导...
在发表的文章中,有一个错误。在方法部分中已经确定了与靶向矢量构建以进行同源重组的描述有关的印刷错误。对材料和方法进行了校正,4.3构造靶向向量以用于胚胎干细胞中的同源重组。这句话先前指出:“同源臂是通过从C57BL/6N小鼠基因组DNA克隆来分离的。在C57BL/6N小鼠ES细胞中生成了针对特定的CD3 G,CD3 D和CD3 E细胞外结构域基因的三个靶向载体。对于CD3 G靶向载体,通过DNA合成将人性化嵌合CD3 G cDNA引入了鼠外显子3中。在3'人源化盒中插入了loxp-抗呼吸毒素的抗性盒。对于CD3 D靶向载体,通过DNA合成将人性化嵌合CD3 D cDNA引入鼠2。在3'的人源化盒中插入了FRT抗抗霉素的耐药盒。对于CD3 E靶向载体,通过

脂肪酸合酶(FASN)抑制剂和甲状腺激素受体β(THRB)激动剂的组合显示
•雄性C57BL/6J Gubra-氨基蛋白 - 纳什(GAN)饮食诱导的肥胖小鼠,具有组织学确认的NAS(≥5)(≥5)和纤维化阶段(F2-F3)(F2-F3),并用TVB-3664(TVB-3664)(替代fasn imbioritor for denifater in demifitor for denifanStat,5 mg/keg)进行治疗(MGL-3196,3 mg/kg,PO,QD)单独或组合6周(每组n = 10-12,Gubra,丹麦)

diO3可预防甲状腺毒性衍生的颅骨...
孕产妇甲状腺功能亢进与出生时先天异常的发生率增加有关,但是尚不清楚这些缺陷中的哪一个是由于持续过量的甲状腺激素过量发育而产生的,哪些依赖于妊娠期,哪些依赖于妊娠阶段,抗肌瘤药物的选择,或抗抑制性药物,或者是不愿意的。为了解决这个问题,我们研究了一个综合发育性甲状腺毒素的小鼠模型,该模型继发于缺乏3型去碘酶(DIO3)。dio3 - / - 小鼠在大多数遗传背景上表现出降低的新生儿生存力,而在C57BL/6背景上表现出围产期致死性。dio3 - / - 小鼠在新生儿期和软骨损失期间表现出严重的生长迟缓。出生后存活的小鼠表现出大脑和颅内畸形,严重的脑积水,choanal闭锁和口感。除了甲状腺毒性心脏,具有隔中间缺陷和稀薄的心室壁,这些异常在胎儿的C57BL/6J dio3 - / - 小鼠中很明显。我们的发现强调了DIO3在开发过程中的保护作用,并支持以下假设:妊娠期间与甲状腺功能亢进相关的人类先天性异常是由临床干预之前的短暂性甲状腺毒性引起的。我们的结果还表明,甲状腺激素参与特发性病理学的病因,包括left裂,Choanal闭锁,Chiari畸形,Kaschin-Beck病以及Temple以及其他颅骨脑膜炎和心脏综合征。