XiaoMi-AI文件搜索系统
World File Search SystemHEK293T

量子硬件上的超跨性汉密尔顿特征状态的实时演变
摘要CRISPR/CAS9系统最初是从原核生物适应性免疫系统中得出的,已作为有效的基因组编辑工具开发。它可以通过可编程SGRNA与靶DNA的特定结合对染色体DNA进行精确的基因操纵,并且具有内切核酸酶活性的CAS9蛋白将在特定位点减少双链断裂。然而,CAS9是哺乳动物细胞中的一种异物,与引入哺乳动物细胞有关的潜在风险尚不完全了解。在这项研究中,我们对HEK293T细胞中的链球菌CAS9(Spycas9)进行了下拉和质谱分析(MS)分析,并表明大多数Cas9-相关蛋白质由MS鉴定的大多数相关蛋白在核中局部局部。有趣的是,我们进一步发现CAS9蛋白包含编码核仁拘留信号(NODS)的序列。与野生型(WT)Cas9相比,CAS9的点突变变体(MCAS9)较小
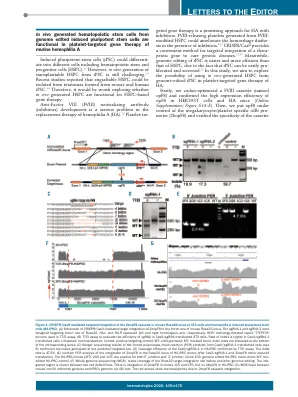
基因组编辑诱导多能干细胞体内产生的造血干细胞可用于小鼠血友病 A 的血小板靶向基因治疗
基因治疗是针对含有抑制剂的HA的一种有前途的方法。由FVIII修饰的HSPC产生的释放FVIII的血小板可以在抑制剂存在的情况下改善出血素质。9-11 CRISPR / Cas9为靶向整合治疗基因以治疗遗传疾病提供了一种便捷的方法。12-14同时,由于iPSC易于增殖和筛选,因此iPSC的基因组编辑比HSPC更容易,效率更高。15在本研究中,我们旨在探索使用基因组编辑的iPSC体内产生的HSPC进行血小板靶向基因治疗HA的可能性。首先,我们对FVIII盒(命名为opF8)进行了密码子优化,并证实了opF8在HEK293T细胞和HA小鼠中的高表达效率(在线补充图S1A-E)。然后,我们将 opF8 置于巨核细胞/血小板特异性 α IIb 启动子 (2bopF8) 的控制之下,并验证了该盒的特异性

Plasmid2MC:高效无细胞重组质粒成高纯度微环 DNA 用于基因组编辑应用
DNA 质粒通常用于在基因组编辑中传递蛋白质和 RNA。然而,与缺乏此类细菌序列的微环 DNA (mcDNA) 相比,它们的细菌成分可能导致失活、细胞毒性和效率降低。现有的将质粒重组到专有细菌菌株内的 mcDNA 中的商业试剂盒劳动密集型,产生的结果不一致,并且通常产生低质量的 mcDNA。为了解决这个问题,我们开发了 Plasmid2MC,这是一种使用 Φ C31 重组的无细胞方法,可有效从常规制备的质粒中切除细菌骨架,而 mcDNA 纯化步骤可消化所有 DNA 杂质并降低内毒素水平。我们展示了 mcDNA 表达 CRISPR-dCas9 在 HEK293T 细胞和小鼠胚胎干细胞中的碱基编辑以及同源性独立的靶向插入 (HITI) 基因组编辑中的应用。该方法易于制备、效率高且 mcDNA 纯度高,使其成为需要细菌无骨架环状 DNA 的应用的宝贵替代方案。

乙酰转移酶p300调节NRF2的稳定性和定位
转录因子 NRF2 在保护细胞免受环境压力和维持细胞稳态方面起着关键作用。乙酰转移酶 p300 是 NRF2 转录复合物的已知组成部分,可促进其转录活性。在本研究中,我们描述了 p300 促进 NRF2 活性的一种新机制。p300 与 NRF2 发生物理相互作用并干扰 NRF2-KEAP1 复合物的形成。特别是,p300 增加了 NRF2 蛋白的丰度和稳定性,从而促进了 NRF2 的核定位。值得注意的是,p300 的乙酰转移酶活性对于 NRF2 的稳定作用是必不可少的。此外,过表达 p300 可保护 HEK293T 细胞免受氧化应激并提高其活力。总之,我们的研究揭示了 p300 与通过调节 NRF2 稳定性来控制 NRF2-KEAP1 信号传导之间的联系,这可能成为适应氧化应激的新型检查点。© 2020 作者。由 Elsevier Inc. 出版。这是一篇根据 CC BY-NC-ND 许可协议开放获取的文章(http://creativecommons.org/licenses/by-nc-nd/4.0/)。

寻找并切割转移 (FiCAT) 哺乳动物基因组工程
虽然存在多种用于小等位基因基因组编辑的技术,但仍然缺乏用于在哺乳动物基因组中靶向整合大 DNA 片段的强大技术。在这里,我们开发了一种基因传递工具 (FiCAT),它结合了 CRISPR-Cas9(发现模块)的精确度和工程化 piggyBac 转座酶(切割和转移模块)的有效载荷转移效率。FiCAT 结合了 Cas9 DNA 扫描和靶向 DNA 的功能以及 piggyBac 供体 DNA 处理和转移能力。PiggyBac 功能域经过工程设计,可提高靶向整合率,同时减少脱靶事件。我们展示了在细胞(人类(Hek293T、K-562)和小鼠(C2C12))和小鼠肝脏体内有效传递和可编程插入小型和大型有效载荷。最后,我们通过生成 394,000 个变体的靶向多样性并进行 4 轮进化,开发出更高效的 FiCAT 版本。在这项工作中,我们开发了一种在哺乳动物基因组中精确、有效地靶向插入多千碱基 DNA 片段的方法。

个体年龄相关 CpG 的表观遗传编辑会影响全基因组的表观遗传衰老状况
PDE4C 中的区域保持稳定超过三个月。此外,表观遗传编辑引发了许多全基因组脱靶效应,这些效应具有高度可重复性,并在其他与年龄相关的 CpG 中富集 - 因此,它们不是随机的脱靶效应,而似乎类似于共同调节的表观遗传旁观者修饰。年龄相关位点的 4C 染色质构象分析显示与旁观者修饰和其他与年龄相关的 CpG 位点的相互作用增加。随后,我们在 HEK293T 和原代 T 细胞中的五个基因组区域多重分析了表观遗传修饰,这些区域在衰老时会变得高甲基化或低甲基化。虽然在年龄低甲基化的 CpG 处进行的表观遗传编辑似乎不太稳定,但它也导致其他与年龄相关的 CpG 处旁观者修饰明显富集。相反,表观遗传时钟往往会在靶向 DNA 甲基化后长达十年内加速,尤其是在高甲基化的 CpG 处。这些结果表明,有针对性的表观基因组编辑可以调节整个表观遗传衰老网络,从而干扰表观遗传时钟。

现场单一构造系统的开发......
摘要:定点 RNA 编辑 (SDRE) 技术在治疗点突变引起的遗传疾病方面具有巨大潜力。我们小组和其他研究人员已经开发出利用作用于 RNA 的腺苷脱氨酶 (ADAR) 和引导 RNA 招募 ADAR 来靶向带有点突变的 RNA 的 SDRE 方法。一般来说,有效的 SDRE 依赖于引入相对于靶基因的大量引导 RNA。然而,对于基因治疗应用来说,实现较大的比例是不可能的。为了实现现实的比例,我们在此开发了一种系统,该系统可以使用由 ADAR 片段组成的融合蛋白和在单个构建体上包含每个基因一个拷贝的质粒载体将相等数量的基因和引导 RNA 引入培养细胞。我们将单个构建体转染到 HEK293T 细胞中并实现了相对较高的效率(高达 42%)。结果表明,当所有三个因素(靶基因、引导 RNA 和 ADAR 酶)的拷贝数相似时,可以实现有效的 SDRE。该方法有望在体内实现高效的基因修复,从而可应用于基因治疗。

Prime编辑允许在负责Duchenne肌肉营养不良的基因中引入特定突变
摘要:从CRISPR/CAS9发现得出的主要编辑技术允许在特定基因中对选定的核苷酸进行修改。我们用它在外显子9、20、35、43、55和61中插入了特定的点突变,该基因肌营养不良蛋白编码为肌营养不良蛋白,该基因在DMD患者中不存在。分别用Prime Editor 2(PE2)和PE3获得了HEK293T细胞中DMD基因的11%和21%所需的突变。三种重复治疗将PE2的特定突变的百分比增加到16%。在单次治疗后,原始的邻接基序(PAM)序列中的额外突变提高了PE3结果至38%。我们还对患者成肌细胞中DMD基因的外显子6中的外显子6中的c.428 g>进行了校正。成肌细胞电穿孔分别显示高达8%和28%的修饰。成肌细胞校正导致通过蛋白质印迹检测到的肌管中肌营养不良蛋白的表达。因此,可以使用序数编辑来校正DMD基因中的点突变。

血清素诱导的 HRP 介导邻近标记抑制
邻近依赖性生物素化与质谱联用可以表征亚细胞蛋白质组。该技术通过揭示亚突触蛋白质网络(例如突触间隙和突触后密度)显著推动了神经科学的发展。在这种详细水平上分析蛋白质对于理解神经元连接和传递的分子机制至关重要。尽管邻近标记最近成功应用于各种神经元类型,但它尚未用于研究血清素系统。在这项研究中,我们发现了血清素对基于辣根过氧化物酶 (HRP) 的生物素化的未报道的抑制机制。我们的结果表明,血清素显着降低 HEK293T 细胞和原代神经元中不同生物素-XX-酪胺 (BxxP) 浓度的生物素化水平,而多巴胺的干扰最小,突出了这种抑制的特异性。为了抵消这种抑制,我们证明了 Dz-PEG(一种通过偶氮偶联反应消耗血清素的芳基重氮化合物)可恢复生物素化效率。无标记定量蛋白质组学证实血清素会抑制生物素化,而 Dz-PEG 可有效逆转这种抑制。这些发现强调了在邻近依赖性生物素化研究中考虑神经递质干扰的重要性,尤其是对于神经科学中细胞类型特异性分析而言。此外,我们还提供了一种缓解这些挑战的潜在策略,从而提高此类研究的准确性和可靠性。

CRISPR-CAS9核糖核蛋白的包装递送加速基因组编辑
图1:用荧光相关光谱(FCS)量化CAS9 RNP核浓度a)工作流的实验示意图,以量化编辑所需的CAS9 RNP核浓度。b)Cas9 rnp或gRNA的扩散时间,在每个细胞中,在Hela细胞中传递,并在24小时(2.0 vs 1.0 ms,p值= 0.0004)时测量每个点,代表用两种组件扩散拟合模拟的单个细胞中平均扩散时间(图。s1)。fcs在MS中提供的扩散时间,每个FCS条件中提供至少两个生物学重复(平均值±SEM)。c)用Cas9 RNP电穿孔的HeLa细胞的FCS分析。Cas9 RNP的核浓度是剂量的函数(每个细胞Cas9)。 每个点表示单个细胞中的浓度。 fcs值在NM中提供,每个FCS条件至少有两个生物学重复(平均值±SEM)。 通过将FCS跟踪与两个组件3D扩散方程拟合(有关详细信息,请参见方法),从而得出了所有浓度值和扩散时间。 d)(左轴)CAS9 RNP与剂量的平均核浓度显示出很强的线性相关性。 (r 2 = 0.96)。 (右轴)由FCS计算出的核浓度值和HeLa核的体积(690μm3)估计的Cas9数量(46)。 e)HELA,U2OS和HEK293T细胞的核浓度的FCS分析。 fcs值在NM中提供,每个FCS条件至少有两个生物学重复(平均值±SEM)。Cas9 RNP的核浓度是剂量的函数(每个细胞Cas9)。每个点表示单个细胞中的浓度。fcs值在NM中提供,每个FCS条件至少有两个生物学重复(平均值±SEM)。通过将FCS跟踪与两个组件3D扩散方程拟合(有关详细信息,请参见方法),从而得出了所有浓度值和扩散时间。d)(左轴)CAS9 RNP与剂量的平均核浓度显示出很强的线性相关性。(r 2 = 0.96)。(右轴)由FCS计算出的核浓度值和HeLa核的体积(690μm3)估计的Cas9数量(46)。e)HELA,U2OS和HEK293T细胞的核浓度的FCS分析。fcs值在NM中提供,每个FCS条件至少有两个生物学重复(平均值±SEM)。在补充表2中报告了FC的精确值,包括实验和生物学重复,平均值和SEM