XiaoMi-AI文件搜索系统
World File Search SystemOncogene

克服胰腺癌的治疗耐药性
致癌作用是由致癌基因 Kirsten Ra Sarcoma (kras) 的驱动突变和肿瘤抑制基因(如 tp53、细胞周期蛋白依赖性蛋白激酶 (cdkn2a)、抗 decapentaplegic 同源物 (smad)-3 和 − 4 的缺失突变逐渐积累引起的 [3] 。乳腺癌 A2 (brca2) 和 A1 (brca1)、brca2 的伴侣和定位器 (palb2)、毛细血管扩张性共济失调突变 (atm)、mutL 同源物 1 (mlh1)、mutS 同源物 2 (msh2) 和 6 (msh6) 中的种系突变也与胰腺癌易感性有关,在 4 – 19 % 的遗传性 PDAC 中发现 [4] 。这些基因改变伴随着胰腺导管细胞内的组织学变化,导致癌前病变(称为胰腺上皮内瘤变 (PanIN))的等级不断提高。

靶向治疗非小细胞肺癌中的致癌基因 KRAS
简单总结:v-Ki-ras2 Kirsten 大鼠肉瘤病毒致癌基因 ( KRAS ) 是 NSCLC 中最常见的驱动因素,靶向致癌 KRAS 是治疗非小细胞肺癌 (NSCLC) 的一大挑战。虽然几种共价 KRAS G12C 抑制剂已成为新型抗 KRAS 疗法,但鉴于 KRAS 突变肿瘤的巨大异质性,仍然需要开发涉及靶向致癌 KRAS 加上其他靶向药物的联合疗法。在这篇综述中,我们总结了致癌 KRAS 驱动的 NSCLC 的生物学和免疫学特征以及突变 KRAS 靶向治疗的临床前和临床证据。我们还讨论了对 KRAS G12C 抑制剂的耐药机制以及克服这种耐药性的可能治疗策略。

克服胰腺癌的耐药性
致癌作用是由癌基因Kirsten RA SARMA(KRAS)中驱动突变的逐渐积累以及肿瘤抑制基因的功能丧失突变引起的,例如TP53,Cyclin-依赖性激酶(CDKN2A),母亲(母亲),针对Decentapplegic filestaplegic同源物(Smad)-3和-3和-4 [3]。BRCA2(PALB2)的合作伙伴和本地化,乳腺癌A2(BRCA2)和A1(BRCA1)中的种系突变,Ataxia telangictia症突变(ATM),MUTL同源1(MLH1),MUTS同源2(MSSH2)和6(MSH2)和6(MSH6)也与Prantis cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer cancer canters cantl of sentoccant也很少相关。这些遗传改变伴随着胰腺导管细胞内的组织学变化,导致

抑制 miMOMP 诱导的 SASP 以对抗与年龄相关的疾病
细胞衰老于 1961 年首次被描述,最初是在正常人类成纤维细胞中观察到的,这些细胞在培养中经过有限次数的分裂后停止增殖。该过程由各种刺激引发,包括氧化应激、染色质修饰和致癌基因激活,其特征是不可逆的细胞周期停滞、对细胞凋亡的抵抗以及复杂的衰老相关分泌表型 (SASP) 的诱导。在过去十年中,新出现的证据将细胞衰老与衰老过程和多种慢性年龄相关疾病联系起来。因此,针对衰老以缓解或延缓与年龄相关的疾病(称为衰老疗法)的研究已经迅速开展。因此,阐明细胞衰老的机制对于提供旨在解决这一状况的实用策略至关重要。

残留宿主细胞 DNA 清除的评估和……
基于病毒载体的基因治疗药物产品可能含有来自生产细胞的残留 DNA。生产通常发生在具有致瘤潜力的连续细胞系中,残留宿主细胞 DNA 具有将致癌基因传播给受体细胞的理论风险。这种风险随着残留 DNA 片段浓度和大小的降低而降低。因此,监测基因治疗产品中残留宿主细胞 DNA 的浓度和大小非常重要。我们评估了在 HEK293T 细胞中制造的工程慢病毒载体生产过程中定量残留宿主细胞 DNA 浓度和大小的方法,并使用这些方法比较了两种市售核酸酶在下游加工步骤中的表现。

casp9作为预后生物标志物和有前途的药物靶标在炎症性乳腺癌中起关键作用
癌症治疗[8]。不良,BCl-2相关的死亡启动子,通过调节细胞周期进展,调节乳腺癌细胞的增殖和肿瘤进展,使乳腺癌细胞对化学疗法敏感[24,25]。SSX2相互作用蛋白(SSX2IP)来调节睾丸和恶性细胞中SSX2的活性[26]。许多研究报告说,SSX2IP作为急性髓样白血病相关的抗原是白血病的潜在免疫疗法靶标[27,28]。还推测SSX2IP在胃癌和肝癌的发展和转移中起重要作用[29]。Znf24起着乳腺癌和胃癌肿瘤发展的负调节剂[30,31]。Znf24也充当癌基因并促进了前列腺癌细胞的EMT [32]。但是,

对雌激素受体-α阳性乳腺癌的潜在多重靶标抑制剂的药物筛查
摘要。背景/目标:雌激素受体α(ERα)拮抗剂是ERα阳性乳腺癌的最常见治疗方法。但是,代偿信号传导有助于对ERα拮抗剂的抗性。因此,为了探索靶向补偿信号传导的潜在药物,我们筛选了多种靶标抑制剂进行乳腺癌治疗。材料和方法:我们试图建立一个基于结构的虚拟筛选模型,该模型可以通过整体细胞存活分析来找到潜在的化合物并测定这些药物的抗癌能力。通过免疫印迹测量下游补偿性磷酸化信号传导。结果:Hamamelitannin和Glucocheirolin均为ERα,磷酸肌醇3-激酶(PI3K)和KRAS原始癌基因,GTPase(KRAS)(KRAS)的命名,它们对雌激素和表皮生长因子触发的增殖具有活性。
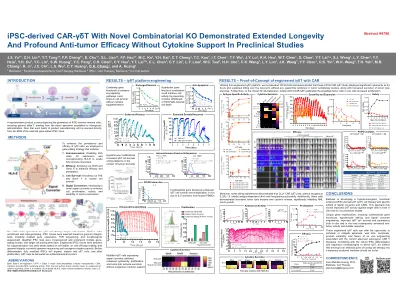
具有新型组合 KO 的 iPSC 衍生 CAR-γδT 在临床前试验中无需细胞因子支持即可表现出更长的寿命和显著的抗肿瘤功效
用于 iγδT 细胞疗法的 GMP 克隆生成始于人类 PBMC。在富集和重编程后,根据基因组完整性测试(包括残留基因表达、TCR 测序和形态学评估)选择 iPSC 克隆。合格的 iPSC 系被冷冻保存并经过多轮基因编辑,每轮之后进行单细胞分选。根据细胞健康、靶向和脱靶编辑以及基因组完整性(通过全基因组测序和致癌基因突变面板)选择工程 iPSC 克隆进行冷冻保存到种子库中。在分化之前,完全改造的 iPSC 将扩增、成熟为 γδT 细胞,增殖后,iγδT 细胞被收获为药品。

抗 CD25 靶向疗法作为免疫调节剂和癌症靶向药物的新兴作用
图 1. 受刺激 T 细胞中的 IL2R 激活途径表示。IL2R 的不同构象会影响其对 IL2 的亲和力(低亲和力 CD25 或高亲和力三聚体受体)。IL2 还可以通过 CD122/CD132 二聚体影响信号传导。此外,形成受体的 CD25 分子是来自相邻细胞(反式)还是同一细胞(顺式)决定了高亲和力异三聚体受体的命名惯例。途径的激活由 Janus 激酶 1 和 3(JAK1 和 JAK3)磷酸化启动,进而刺激 STAT5 二聚化,或磷酸肌醇 3 激酶 (PI3K) 和大鼠肉瘤病毒致癌基因同源物 (Ras) 途径,最终磷酸化效应激酶 p70 S6K 和 MAPK。

正常生理学中的细胞衰老
世界人口的老化对理解衰老过程,制定策略和干预措施以延长健康寿命的兴趣加剧了兴趣。细胞衰老,当细胞在体外细胞增殖后或对绝对应激或癌基因表达反应后不可逆生长时(1,2)在AG的表型和年龄相关疾病中起作用(1)。越来越多的证据表明,衰老细胞也具有必不可少的生理功能,例如在肿瘤抑制,发育,伤口愈合,组织重塑,再生和脉管系统中。这就提出了有关衰老细胞类型之间的相似性和差异以及它们在体内平衡和病理学中的作用的重要问题,并在治疗方面靶向它们时会产生更多的挑战。