XiaoMi-AI文件搜索系统
World File Search System不饱和

使用自电位表面 (SPS) 进行洞穴探测......
1 degéomagnétisme,瑞士大学,瑞士,marcus.gurk@unine.ch 2中心D'Hydrogéologie,瑞士大学,瑞士大学,弗兰克(Frank.bosch.bosch.bosch.bosch.bosch@unine.ch exprient for Selferation for Selferation for Selferation for Selferation forefface facee)火山区。前提是满足了两个条件,他们在实验上发现了负自力(SP)异常的范围与不饱和区域的厚度之间的线性关系。第一个条件是不饱和区域的电阻率与底层和水饱和区的电阻率之间的强对比度。第二条件是不饱和区域的同质性。SP地图的定性解释表明,最大负值的线对应于排水轴和两个分水岭之间边界的最小负值线(Jackson&Kauahikaua(1987))。我们期望类似的条件在碳酸盐含水层中有效。尤其是在karstic洞穴中,空气层产生的电阻率对比必须很重要,因此SPS技术可用于检测这些结构。是由这一假设的动机,与水文地质学中心Neuchâtel(Chyn)进行了实验。2调查区域实验实验是在Vers-Chez-Le-Brandt(Bosch&Gurk,2000年)的洞穴上进行的,该实验是在法国附近的瑞士Jura Mountains,CantonNeuchâtel的折叠式石灰石中进行的(图。1)。在该地区中生代石灰石和泥浆中,被薄薄的季节沉积物覆盖。洞穴的发展为腔/阿格维亚上喀布尔的石灰石的方向约为N140°(图2),长度约为260m。该序列的泥土层允许开发一条小的地下河。构造特征(例如断层)在洞穴内可见(Müller,1981)。这些罢工方向与瑞士折叠的jura中控制压裂和凸出的局部应力参数(主剪切= N0°,σ1= N130°-N150°,σ2= N40°-N50°)。

共焦拉曼显微镜的食物分析
在一个相关的例子中,拉曼成像用于比较两种黄油产物,以研究其不同扩展能力的基础化学差异。通过沿z轴相结合在连续的焦平面上获得的2D图像来产生正常黄油和更可扩展产物的单个3D拉曼图像(图5a,b)。这两种产品显然是预期的油脂层。在可撒黄油中的水含量高,水的水与更坚固的脂肪相比,水含量更大。化学物质在脂肪阶段中的分化通过比较其拉曼光谱而变成证明(图5C)。 每种产品都包含不同类型的脂肪和油。 脂肪的同意受不饱和脂肪酸的量以及其他参数的影响。 可以通过1655 cm -1 的C = C拉伸模式的比率比较脂肪的不饱和度5C)。每种产品都包含不同类型的脂肪和油。脂肪的同意受不饱和脂肪酸的量以及其他参数的影响。可以通过1655 cm -1

抗氧化剂生产的可持续策略
摘要:不饱和二酰基甘油是一类抗氧化剂化合物,具有积极影响人类健康的可能性。他们通过根治性的清道夫活动能够打击氧化应激的能力强调了其在预防和治疗策略的背景下的意义。在本文中,我们强调了Anabaena flos-aquae作为不饱和单甘油和二酰基甘油的生产国的作用,然后证明其甲醇提取物的抗氧化活性,其主要成分是各种乙酰甘油类似物。使用可持续策略揭示了这一发现,其中一种菌株在微观中,许多化合物(OSMAC)培养与生物信息学方法结合,以使用分子网络分析来分析大量质谱数据数据集。此策略减少了时间和成本,避免了纯净的纯净步骤,并获得有关提取物代谢组成的信息数据。这项研究强调了Anabaena作为新型生物活性化合物的可持续和绿色来源的作用。
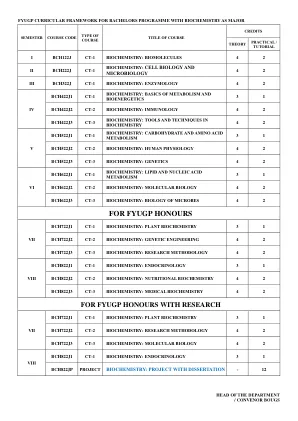
for fyugp荣誉
脂肪酸的简介,分类,命名法,结构和特性。饱和和不饱和脂肪酸。必需脂肪酸,化学特性和脂肪的表征 - 水解,皂化值,雷克特 - 梅塞尔数,碘数,脂肪的酸味,脂肪的酸味,三酰基甘油和胆固醇,磷脂和鞘脂的结构和功能。前列腺素和类固醇激素的合成。

HJC PIM(高级集成矩阵)技术
HJC PIM Shell 由多种高科技面料混合而成。虽然目前大多数头盔采用的是普通不饱和聚酯树脂和普通玻璃纤维,但我们结合了尽可能高水平的先进材料。这种先进优质复合材料的使用,得益于其坚固而柔韧的斜纹编织加固,可产生极高的冲击吸收率和出色的轻盈度。

反刍动物营养中脂肪酸的封装...
近年来,生产饲料用保护性脂肪补充剂的方法得到了很大的发展。作为防止不饱和脂肪氧化的一种方法,食品工业研究人员使用包封来减少不愉快的气味和味道,并作为保护不饱和脂肪的有效方法。包封过程涉及将目标物质覆盖或捕获在另一种物质或系统中。同样,食品中的维生素和微量营养素化合物不会长时间保持稳定,并且容易分解,这取决于化学结构、食品基质特性、处理参数和储存条件。因此,包封可以防止这些化合物被破坏,直到它们被转移到正确的位置或减缓分解过程(如氧化或水解)。这一概念可以扩展到脂质(油和脂肪)。目前,乳液喷雾干燥是精细油微涂层的最常见方法。最近发现,与喷雾干燥相比,团聚形成方法可以产生更稳定的微涂层,油含量更高。可生物降解的聚合物作为包封材料引起了广泛关注。微囊化脂质可以提高反刍动物的肉和奶的品质。

微生物风险评估工具文档报告
图2:现场废水管理系统(OWMS)和地下水抽象点之间微生物去除的组成部分。1)处理厂内的微生物还原,2)土地申请系统内的微生物还原(LAS),3)在不饱和(vadose)区域内的微生物还原,4)在饱和区内的微生物还原。注意:抽象井直接下降了LA的梯度。红色箭头给出了流动的方向,并预测微生物浓度从OWMS良好到抽象的降低。............................................................................. 37

光子响应性木质素片段基于可切换粘合剂
易于拆卸和可重复使用的粘合剂作为一次性粘合剂的替代品具有吸引力,可减少浪费并促进再利用,回收或什至升级选项。木质素是纸 - 羽状产业的第二大聚合物和副产品,用于设计一种新颖的,高度可调的可逆聚合物粘合剂。采用的方法是利用P-羟基霉素酸在这项工作中使用木质素氧化化合物合成的P-羟基霉素酸结构的α,β-不饱和酯部分的光子响应特性,并使用木质素氧化化合物合成并修饰以可耐可可逆的粘附切换。可逆性是通过紫外线的暴露来实现的,紫外线裂解最初由酯的α,β-不饱和键形成的共价环丁烷环,从而使材料变软并易于分离。可以通过弹性链接以提供重新功能来再次建立原始聚合物结构。引入了实验方法(DOE)方法的设计,以优化重要变量,以实现粘合剂的最佳剪切强度。各种结构方面的效果显示了满足财产要求的结构的高可调节性。可再生资源的聚合物粘合剂的设计策略,以及本工作中描述的结构 - 属性分析机制,可以实施以设计基于生物的新型和可重复使用的粘合剂。

提高你的自然生育能力
• 富含植物性食物,包括水果、蔬菜、豆类、坚果和种子 • 富含单不饱和脂肪,如橄榄油 • 饱和脂肪和反式脂肪含量低 • 吃少量家禽和极少的红肉 • 包括鱼,尤其是油性鱼 • 吃少量乳制品——主要是天然酸奶和少量奶酪 • 糖分低,甜味主要来自新鲜水果和干果 • 吃少量家禽和极少的红肉 最好避免食用超加工食品和饮料。

带有 ReliaTel™ 控件的 LonTalk® 通信接口
科学研究表明,某些人造化学物质在释放到大气中时会影响地球自然形成的平流层臭氧层。具体来说,已确定的几种可能影响臭氧层的化学物质是含氯、氟和碳 (CFC) 的制冷剂以及含氢、氯、氟和碳 (HCFC) 的制冷剂。并非所有含有这些化合物的制冷剂都对环境具有相同的潜在影响。特灵倡导负责任地处理所有制冷剂,包括 CFC 和 HCFC 的行业替代品,例如饱和或不饱和 HFC 和 HCFC。