XiaoMi-AI文件搜索系统
World File Search System去极化

延迟 GABA 转移会间接影响发育中的海马体的膜特性
在出生后的前两周,啮齿动物的神经元内氯离子浓度逐渐下降,导致 GABA 反应从去极化转变为高极化。在神经发育障碍的啮齿动物模型和人类患者中,出生后的 GABA 转变会延迟,但 GABA 转变延迟对发育中大脑的影响仍不清楚。在这里,我们通过用氯离子输出蛋白 KCC2 的特异性抑制剂 VU0463271 处理 6 至 7 日龄小鼠的器官型海马培养物 1 周,研究了出生后 GABA 转变延迟对网络发育的直接和间接影响。我们证实了 VU 治疗延迟了 GABA 转变并使 GABA 信号去极化直到 DIV9。我们发现 VU 治疗后 DIV9 时的兴奋性和抑制性突触的结构和功能发育没有受到影响。与之前的研究一致,我们观察到 GABA 信号在对照组和 VU 处理的出生后切片中已经受到抑制。令人惊讶的是,在 VU 治疗结束 14 天后(DIV21),我们观察到 CA1 锥体细胞中自发抑制性突触后电流的频率增加,而兴奋性电流没有改变。突触数量和释放概率不受影响。我们发现,与对照切片相比,放射层中以树突为靶向的中间神经元具有升高的静息膜电位,而锥体细胞的兴奋性较低。我们的结果表明,去极化 GABA 信号不会促进 P7 后的突触形成,并表明出生后细胞内氯离子水平以细胞特异性的方式间接影响膜特性。

BIO 105生物心理学
去极化通过阈值创造了动作电位。钠通道打开,并使Na+进入神经元。当电势到达0钠通道时。然后钾通道打开,钾降低。这将神经元返回到静止电位。这种化学物质的交换沿轴突延伸,动作电位通过轴突传播。

使用张量网络解码器对泡利噪声进行量子编码的随机 Clifford 电路
其中 p I + p X + p Y + p Z = 1。我们主要考虑去极化噪声的情况 p X = p Y = p Z = p / 3,p I = (1 − p )。▶ 众所周知 1 使用随机 Clifford 单位向量进行编码,可以实现称为哈希界限的速率

跨...
摘要:在电缆中的绝缘层的交联聚乙烯(XLPE)的广泛使用可能归因于其出色的机械和介电性能。为了定量评估热老化后XLPE的绝缘状态,建立了加速的热老化实验平台。极化和去极化电流(PDC)以及在不同老化持续时间下XLPE绝缘裂纹时的伸长率。XLPE绝缘状态取决于断裂保留率(ER%)的伸长率。基于扩展的Debye模型,本文提出了稳定的松弛电荷数量和0.1 Hz的耗散因子,以评估XLPE的绝缘状态。结果表明,XLPE绝缘的ER%随着衰老程度的增长而降低。XLPE绝缘的极化和去极化电流将随着热老化而明显增加。电导率和陷阱水平密度也将增加。扩展Debye模型的分支数量增加,并出现新的极化类型。在本文提出的0.1 Hz处的稳定的松弛电荷量和耗散因子与XLPE绝缘的ER%具有良好的拟合关系,可以有效地评估XLPE绝缘的热老化状态。

量子局部差分隐私的优化机制
集中式差分隐私已成功应用于量子计算和信息处理,以保护隐私并避免相邻量子态之间连接中的泄漏。因此,量子局部差分隐私 (QLDP) 已被新提出以保护量子数据隐私,类似于所有状态都被视为相邻状态的经典场景。然而,QLDP 框架的探索仍处于早期阶段,主要是概念性的,这对其在保护量子态隐私方面的实际实施提出了挑战。本文对 QLDP 进行了全面的算法探索,以建立一个实用且可行的 QLDP 框架来保护量子态隐私。QLDP 使用参数 ε 来管理隐私泄漏并确保单个量子态的隐私。对于任何量子机制,QLDP 值 ε 的优化(表示为 ε ∗ )都是一个优化问题。结果表明,量子噪声的引入可以提供与经典场景类似的隐私保护,量子去极化噪声被确定为 QLDP 框架内的最佳单元私有化机制。单元机制代表了一组多样化的量子机制,涵盖了经常使用的量子噪声类型。量子去极化噪声优化了保真度和迹线距离效用,这是量子计算和信息领域的关键指标,可以看作是经典随机响应方法的量子对应物。此外,提出了一个组合定理,用于将 QLDP 框架应用于分布式(空间分离)量子系统,确保有效性(QLDP 值的加性),而不管状态的独立性、经典相关性或纠缠(量子相关性)。该研究进一步通过分析和数值实验方法探讨了不同量子噪声机制(包括单元和非单元量子噪声机制)之间效用和隐私之间的权衡。同时,这突出了 QLDP 框架中量子去极化噪声的优化。

噪声量子通道中的Qudit状态
技术和理论进步使Qudit国家在量子信息和组合中必不可少。量子算法代表了现代量子信息理论领域中的一个突出应用,为计算加速度提供了经典系统不可能实现的潜力。一种实现量子算法的著名方法涉及创建特定类型的异常纠缠的图形状态。超图状态,也称为多部分纠缠状态或高阶纠缠状态,是量子状态,它们将纠缠概念扩展到钟形状态或图形状态中通常发现的成对相关性之外。他们提供了一个平台来概括最初针对Qubit状态的想法。因此,例如,Qudit状态已在量子传送[1-3],量子计算[4 - 6],量子步行[7 - 9]和量子状态转移[10-12]中发现了应用。量子系统始终受到与环境环境相互作用的噪声的影响[13]。因此,对在嘈杂条件下进化的Qudit国家动态的研究是一个相关问题,我们在这里进行了研究。Qudits是Qubits的较高维度概括,在量子科学和技术的几个领域中变得越来越重要[14,15]。噪声在任何物理系统中总是不可避免的现象。特别是量子噪声具有非常特殊的特征,其效果通过非可逆操作员表征。在本文中,我们专注于研究噪声如何影响量子状态。为了研究噪声对状态的影响,应了解相应的量子通道的特征。量子通道由适当的kraus操作员表示。保真度是对此有用的诊断。我们研究的量子通道是dit-Flip噪声,相位翻转噪声,DIT相相位噪声,去极化噪声,ADC(非马克维亚噪声),非马克维亚倾向噪声和非马克维亚去极化噪声[16,17]。这些通道最初被定义为适用于Qubit。dit-Flip噪声,相位翻转噪声,DIT相相翻噪声和去极化噪声被推广到[3]中的Qudit状态。遵循此方向,我们将Qudits上的ADC(非马尔可夫噪声),非马克维亚式Dephasing和非Markovian去极化噪声进行了推广。针对这些通道中的每个通道计算了原始状态和最终状态之间的忠诚度的分析表达。这有助于根据量子状态评估噪声的影响。连贯性是大多数
![arxiv:2006.16223v3 [Quant-PH] 2021年10月11日](/simg/9\979ee1ca449c8d10adf853c7375e71d39bdb46ce.webp)
arxiv:2006.16223v3 [Quant-PH] 2021年10月11日
摘要最近,我们发现了几种量子算法的候选者,这些算法可以在近期设备中实现,以估算给定量子状态的振幅,这是各种计算任务(例如蒙特卡洛方法)中的核心子例程。这些算法之一是基于并行化量子电路的最大似然估计。在本文中,我们扩展了此方法,使其结合了逼真的噪声效果,然后在超导IBM量子设备上进行实验演示。假设去极化噪声的模型构建最大似然估计器。然后,我们将问题提出为两参数估计问题,相对于目标振幅参数和噪声参数。特别是我们表明存在异常的目标值,其中Fisher信息矩阵变成退化,因此即使通过增加振幅膨胀的数量也无法改善估计误差。实验证明表明,提出的最大似然估计器在查询数量中实现了量子加速,尽管估计误差由于噪声而饱和。估计误差的饱和值与该理论一致,这意味着去极化噪声模型的有效性,从而使我们能够预测量子计算机中硬件组件(尤其是门误)的基本要求(尤其是栅极误差),以实现幅度估计任务中的量子加速。

支持信息:受支持的纳米颗粒的等离子间在纳米级上揭示了粘附:对介质的金属的影响
培训该案。一旦计算出截短物体的确切极性i Z,5,10,16,23 25复杂的效果去极化因子就可以从与等效自由式球形相对应的eq S1中得出。在图S6中绘制了e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e s6 s6和具有代表性纵横比r = 0的截短颗粒。5、1、2、4和给定的t r值(主要文本中的truncation参数de ned)。对于粒子的形状不太接近全球(r>1。5),α由偏振电荷类似于偶极的模式主导。它们的效果去极化因子几乎是恒定接近共振的(请参阅图S6中的仪表板上的垂直线),它们的行为实际上是振荡器。

癫痫的基本机制
第3章癫痫的基本机制John G.R.牛津大学癫痫发作的杰弗里斯药理学系通常涉及神经元的过度解雇和同步。这打断了所涉及的大脑部分的正常工作,从而导致特定类型的癫痫类型的临床症状和符号学。本章将概述癫痫放电的基本机制,特别是在局灶性癫痫的细胞电生理学方面。它将概述阐明癫痫发作期间“超同步”神经元活动的概念的最新进展。局灶性癫痫活性局灶性癫痫发生在新皮层和边缘结构中,包括海马和杏仁核。在一系列实验模型上进行的工作产生了有关简短(约100 ms)癫痫事件的详细理论,该事件类似于在具有局灶性癫痫的人EEG中经常发现的“间歇性尖峰”。实验性间歇放电的特征是突然在当地大多数神经元中同步发生的“阵发性”去极化移位(PDSS)。这些是大型去极化,即2040mV,这使神经元燃烧了快速的动作电位。PDS具有巨大的兴奋性突触后电位(EPSP)的特性,并且取决于谷氨酸,这是大脑中主要的兴奋性突触发射机。这个巨大的EPSP是由同一人群中许多其他神经元的同时激发驱动的。这种连接的概率可以很低。例如,海马中的随机选择的锥体细胞的2%之间的〜12%。PDS还取决于神经元的soma树突区域的内在特性,例如电压 - 敏感的钙通道可以产生缓慢的去极化,从而驱动多个快速(钠通道)动作电位。在许多实验模型上的结合实验和理论工作表明,以下特征足以用于这种癫痫发射:兴奋性(通常是金字塔)神经元必须使连接到突触网络。由于单个突触的特性和/或由于突触前神经元的发射模式(由于电压敏感性的去极化通道引起的爆发爆发意味着突触电位可以汇总)。本质上,神经元需要很有可能将其突触后靶标超过阈值。神经元的种群必须足够大(“最小骨料”类似于核裂变炸弹的临界质量)。此最低骨料允许神经元与几个突触中的几乎所有人群中的所有其他人建立联系,从而使一小部分神经元的活动在适当的条件下可以非常迅速地通过人群传播。不同的联系意味着神经元种群是在近距离进展中募集的。在实验模型中,最小癫痫骨料可以低至10002000神经元,但在人类癫痫灶中可能更大。
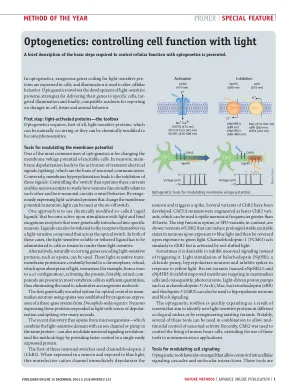
光遗传学:用光控制细胞功能
调节膜电位的工具 光遗传学最常见的用途之一是改变可兴奋细胞的膜电位。在神经元中,膜去极化会导致瞬态电信号(脉冲)的激活,这是神经元通讯的基础。相反,膜超极化会导致这些信号的抑制。控制操作这些电流的“开关”使神经科学家能够研究神经元在功能上如何相互关联以及神经元回路如何控制行为。通过外源表达改变神经元膜电位的光激活蛋白,光可以用作开关。一种方法是使用化学修饰的所谓“笼状配体”,这些配体在光刺激下变得活跃并与通过基因引入特定神经元的外源性受体结合。配体也可以通过充当光开关的光敏化合物与受体本身相连。在这两种情况下,都必须将光敏的可溶性或束缚配体注入细胞或组织,使它们对光敏感。或者,可以使用编码光敏蛋白(如视蛋白)的天然基因。这些光敏跨膜蛋白与发色团视网膜共价结合,视网膜吸收光后发生异构化(例如,从反式变为顺式构型),从而激活蛋白质。值得注意的是,视网膜化合物在大多数脊椎动物细胞中含量充足,因此无需注入外源分子。第一个利用视蛋白进行哺乳动物神经元光学控制的遗传编码系统是通过外源表达果蝇的三基因系统建立的。表达这些蛋白质的神经元对光的反应是数秒内的去极化和尖峰波。最近发现,微生物中的视蛋白(将光敏域与同一蛋白质中的离子通道或泵相结合)也可以调节神经元信号,通过在单个易于表达的蛋白质中提供更快的控制,彻底改变了该方法。这些神经元开关中的第一个使用了通道视紫红质-2 (ChR2)。当在神经元中表达并暴露于蓝光时,这种非选择性阳离子通道会立即使神经元去极化