XiaoMi-AI文件搜索系统
World File Search System腺苷

腺苷脱氨酶缺乏
参考•Blackburn MR,Thompson LF。 腺苷脱氨酶缺乏症:从罕见的免疫缺陷的研究中进行的意外抗原。 J immunol。 2012 Feb1; 188(3):933-5。 doi:10.4049/jimmunol.1103519。 没有抽象可用。 引用于PubMed(https://pubmed.ncbi.nlm.nih.gov/22262755)或PubMed Central上的免费文章(https://wwwwwww.ncbi.nlm.nih.nih.nih.gov/pmc/articles/pmc/articles/ppmc3341658/) Slominska E,Bohynikova N,Bernat-Sitarz K,Bernatowska E,Wolska-Kusnierz B,Kalwak K,Kalwak K,Koltan S,Dabrowska A,Gozdzik J,USSOWICZ J,PAC M. PAC M.具有腺苷Deaminase Deaminase Deficiedicrecity ReficienceReciedrelecipedreled Childs in Polor Polor Polor。 前疫苗。 2023 JAN 6; 13:1058623。 doi:10.3389/fimmu.2022.1058623。 Ecollection2022。 引用PubMed(https://ww w.ncbi.nlm.nih.gov/pubmed/36685585)•Grunebaum E,Booth C,Cuvelier GDE,Loves R,Aiuti A,Aiuti A,Kohn DB。 腺苷脱氨酶缺陷的更新管理指南。 J ALLERGY CLIN IMMUNOLPRACT。 2023 Jun; 11(6):1665-1675。 doi:10.1016/j.jaip.2023.01.032。 EPUB 2023 FEB1。 引用PubMed(https://www.ncbi.nlm.nih.gov/pubmed/36736 952)•Hershfield M,Tarrant T.腺苷脱甲酶缺乏症。 2006年10月3日[更新2024 3月7日]。 in:Adam MP,Feldman J,Mirzaa GM,Pagon RA,Wallace SE,Amemiya A,编辑。 genereviews(r)[Internet]。 西雅图(WA):西雅图大学的大学; 1993-2025。 Curr Opin Immunol。 2003年10月; 15(5):571-7。 doi:10。 1016/S0952-7915(03)00104-3。 EUR J Immunol。 2005 JAN; 35(1):25-30。DOI:10.1002/eji.200425738。参考•Blackburn MR,Thompson LF。腺苷脱氨酶缺乏症:从罕见的免疫缺陷的研究中进行的意外抗原。J immunol。 2012 Feb1; 188(3):933-5。 doi:10.4049/jimmunol.1103519。 没有抽象可用。 引用于PubMed(https://pubmed.ncbi.nlm.nih.gov/22262755)或PubMed Central上的免费文章(https://wwwwwww.ncbi.nlm.nih.nih.nih.gov/pmc/articles/pmc/articles/ppmc3341658/) Slominska E,Bohynikova N,Bernat-Sitarz K,Bernatowska E,Wolska-Kusnierz B,Kalwak K,Kalwak K,Koltan S,Dabrowska A,Gozdzik J,USSOWICZ J,PAC M. PAC M.具有腺苷Deaminase Deaminase Deficiedicrecity ReficienceReciedrelecipedreled Childs in Polor Polor Polor。 前疫苗。 2023 JAN 6; 13:1058623。 doi:10.3389/fimmu.2022.1058623。 Ecollection2022。 引用PubMed(https://ww w.ncbi.nlm.nih.gov/pubmed/36685585)•Grunebaum E,Booth C,Cuvelier GDE,Loves R,Aiuti A,Aiuti A,Kohn DB。 腺苷脱氨酶缺陷的更新管理指南。 J ALLERGY CLIN IMMUNOLPRACT。 2023 Jun; 11(6):1665-1675。 doi:10.1016/j.jaip.2023.01.032。 EPUB 2023 FEB1。 引用PubMed(https://www.ncbi.nlm.nih.gov/pubmed/36736 952)•Hershfield M,Tarrant T.腺苷脱甲酶缺乏症。 2006年10月3日[更新2024 3月7日]。 in:Adam MP,Feldman J,Mirzaa GM,Pagon RA,Wallace SE,Amemiya A,编辑。 genereviews(r)[Internet]。 西雅图(WA):西雅图大学的大学; 1993-2025。 Curr Opin Immunol。 2003年10月; 15(5):571-7。 doi:10。 1016/S0952-7915(03)00104-3。 EUR J Immunol。 2005 JAN; 35(1):25-30。DOI:10.1002/eji.200425738。J immunol。2012 Feb1; 188(3):933-5。 doi:10.4049/jimmunol.1103519。没有抽象可用。引用于PubMed(https://pubmed.ncbi.nlm.nih.gov/22262755)或PubMed Central上的免费文章(https://wwwwwww.ncbi.nlm.nih.nih.nih.gov/pmc/articles/pmc/articles/ppmc3341658/) Slominska E,Bohynikova N,Bernat-Sitarz K,Bernatowska E,Wolska-Kusnierz B,Kalwak K,Kalwak K,Koltan S,Dabrowska A,Gozdzik J,USSOWICZ J,PAC M. PAC M.具有腺苷Deaminase Deaminase Deficiedicrecity ReficienceReciedrelecipedreled Childs in Polor Polor Polor。前疫苗。2023 JAN 6; 13:1058623。 doi:10.3389/fimmu.2022.1058623。Ecollection2022。引用PubMed(https://ww w.ncbi.nlm.nih.gov/pubmed/36685585)•Grunebaum E,Booth C,Cuvelier GDE,Loves R,Aiuti A,Aiuti A,Kohn DB。腺苷脱氨酶缺陷的更新管理指南。J ALLERGY CLIN IMMUNOLPRACT。2023 Jun; 11(6):1665-1675。 doi:10.1016/j.jaip.2023.01.032。EPUB 2023 FEB1。引用PubMed(https://www.ncbi.nlm.nih.gov/pubmed/36736 952)•Hershfield M,Tarrant T.腺苷脱甲酶缺乏症。2006年10月3日[更新2024 3月7日]。in:Adam MP,Feldman J,Mirzaa GM,Pagon RA,Wallace SE,Amemiya A,编辑。genereviews(r)[Internet]。西雅图(WA):西雅图大学的大学; 1993-2025。 Curr Opin Immunol。 2003年10月; 15(5):571-7。 doi:10。 1016/S0952-7915(03)00104-3。 EUR J Immunol。 2005 JAN; 35(1):25-30。DOI:10.1002/eji.200425738。西雅图(WA):西雅图大学的大学; 1993-2025。Curr Opin Immunol。2003年10月; 15(5):571-7。 doi:10。1016/S0952-7915(03)00104-3。EUR J Immunol。 2005 JAN; 35(1):25-30。DOI:10.1002/eji.200425738。EUR J Immunol。2005 JAN; 35(1):25-30。DOI:10.1002/eji.200425738。2005 JAN; 35(1):25-30。DOI:10.1002/eji.200425738。可从http://www.ncbi.nlm.nih.gov/books/ nbk1483/PubMed上获得(https://www.ncbi.nlm.nih.gov/pubmed/20301656)•Hershfield MS。基因型是腺苷酸酶缺乏症中表型的重要决定因素。引用PubMed(https://pubmed.ncbi.nlm.nih.gov/14499267)•Hershfield MS。对腺苷受体介导的免疫抑制和腺苷在引起与腺苷脱氨酶缺乏相关的免疫缺陷中的作用的新见解。引用于PubMed(https://pubmed.ncbi.nlm.nih.go v/15580654)•Nofech-Mozes Y,Blaser SI,Kobayashi J,Grunebaum E,Grunebaum E,Roifman CM。腺苷脱氨酶缺乏症患者的神经学性稳定性。Pediatr Neurol.2007 9月; 37(3):218-21。 doi:10.1016/j.pediatrneurol.2007.03.011。引用于PubMed(https://pubmed.ncbi.nlm.nih.gov/17765813)•nyhan wl。嘌呤和嘧啶代谢的疾病。mol Genet Metab。2005SEP-OCT; 86(1-2):25-33。 doi:10.1016/j.ymgme.2005.07.027。引用于PubMed(https://pubmed.ncbi.nlm.nih.gov/16176880)

回顾腺苷脱氨酶2
腺苷脱氨酶2(ADA2)缺乏是一种常染色体遗传遗传遗传的自身炎症疾病,由ADA2基因的功能丧失突变引起。尽管发病机理涉及促炎性细胞因子的产生,例如肿瘤坏死因子(TNF) - α和中性粒细胞外陷阱形成的失调,而导致的胞外腺苷的积累过多,但仍需要澄清的始终澄清,因此中性粒细胞陷阱形成的产生,以及中性粒细胞外陷阱形成的失调。除了最初描述的与血管炎相关的症状外,血液学,免疫学和自身炎症症状现在已经得到充分认可。诊断是通过证明ADA2的致病变异的,双重功能丧失和低血浆ADA2催化活性的识别。目前,TNF-α抑制剂是控制血管炎表现和预防中风的选择。然而,在出现严重血液学发现的患者中,TNF-α抑制剂不是选择的治疗方法,并且在某些情况下已证明造血干细胞移植已成功。重组ADA2蛋白和基因疗法是未来有希望的治疗方式。总而言之,ADA2缺乏症具有广泛的表型,应在不同临床情况的鉴别诊断中考虑。在这篇综述中,我们总结了ADA2缺乏症和可用治疗方案的疾病表现。

针对 A3 腺苷受体的药物
摘要:A3 腺苷受体 (A3AR) 在病理性人类细胞中过度表达。Piclidenoson 和 namodenoson 是具有高亲和力和选择性的 A3AR 激动剂。两者均通过涉及 Wnt 和 NF- κ B 信号通路失调的分子机制诱导癌症和炎症细胞凋亡。我们公司进行了 I 期研究,证明了这两种分子的安全性。在银屑病患者的 II 期研究中,piclidenoson 是安全的,并显示出显著改善皮肤病变的疗效。Namodenoson 目前正在开发用于治疗肝癌,在患有晚期肝病且 Child-Pugh B 评分为 7 的患者中观察到总生存期延长。针对该患者群体的关键 III 期研究已获得 FDA 和 EMA 批准,目前正在进行中。 Namodenoson 还正在开发用于治疗非酒精性脂肪性肝炎 (NASH)。IIa 期研究已成功完成,表明 Namodenoson 具有抗炎、抗纤维化和抗脂肪变性作用。NASH 的 IIb 期研究目前正在招募患者。总之,A3AR 激动剂是处于临床开发后期的有希望的候选药物,并在其目标适应症中表现出安全性和有效性。

多聚腺苷的动力学和静态分析
作者分支机构:美国宾夕法尼亚州宾夕法尼亚州宾夕法尼亚大学佩雷尔曼医学院放射学系1。2宾夕法尼亚州宾夕法尼亚大学佩雷尔曼医学院妇科和肿瘤学系OBGYN系。 3辐射肿瘤学系,辐射肿瘤学系3,美国德克萨斯州德克萨斯大学医学博士安德森癌症中心,美国德克萨斯州休斯敦 *通讯作者:宾夕法尼亚州宾夕法尼亚州博士学位160A约翰·摩根大楼360年3620 Hamilton Walk PAREDELPHIA,汉密尔顿步行PA 19104电话:(215)573-603-603-603-603-603-603-603-603-603--603--603--603--603--603--603--603--603--603--603--603--603--603--603--603--603--603--603--63-6016: robdoot@pennmedicine.upenn.edu https://orcid.org/000000-0003-1747-239X第一作者:Anthony J. 宾夕法尼亚州宾夕法尼亚州宾夕法尼亚州宾夕法尼亚州的年轻大学1910年约翰·摩根大厦(John Morgan Walk),宾夕法尼亚州19104电话:(215)746-0039传真:(215)573-3880电子邮件:anthony.youn.youn.youn.young@pennmedicine.upenn.upen.upen.upen.upen.upen.edu word Count:4990 Financial Supports:4990 Fancial Supports:RD DAS NIDAS(k01)。 ll希望基金会的万花筒和玛莎·里夫金基金会(Marsha Rivkin Foundation)支持。 AP由KL2TR001879支持。 研究还由国家研究资源中心和国家卫生研究院(UL1TR000003)的国家研究中心和宾夕法尼亚大学转化医学和治疗学研究所,宾夕法尼亚州放射科学系和NCI(P30CA016520)赞助。 在卵巢癌中运行标题18 f-ftt目标PARP-12宾夕法尼亚州宾夕法尼亚大学佩雷尔曼医学院妇科和肿瘤学系OBGYN系。3辐射肿瘤学系,辐射肿瘤学系3,美国德克萨斯州德克萨斯大学医学博士安德森癌症中心,美国德克萨斯州休斯敦 *通讯作者:宾夕法尼亚州宾夕法尼亚州博士学位160A约翰·摩根大楼360年3620 Hamilton Walk PAREDELPHIA,汉密尔顿步行PA 19104电话:(215)573-603-603-603-603-603-603-603-603-603--603--603--603--603--603--603--603--603--603--603--603--603--603--603--603--603--603--603--63-6016: robdoot@pennmedicine.upenn.edu https://orcid.org/000000-0003-1747-239X第一作者:Anthony J.宾夕法尼亚州宾夕法尼亚州宾夕法尼亚州宾夕法尼亚州的年轻大学1910年约翰·摩根大厦(John Morgan Walk),宾夕法尼亚州19104电话:(215)746-0039传真:(215)573-3880电子邮件:anthony.youn.youn.youn.young@pennmedicine.upenn.upen.upen.upen.upen.upen.edu word Count:4990 Financial Supports:4990 Fancial Supports:RD DAS NIDAS(k01)。ll希望基金会的万花筒和玛莎·里夫金基金会(Marsha Rivkin Foundation)支持。AP由KL2TR001879支持。研究还由国家研究资源中心和国家卫生研究院(UL1TR000003)的国家研究中心和宾夕法尼亚大学转化医学和治疗学研究所,宾夕法尼亚州放射科学系和NCI(P30CA016520)赞助。在卵巢癌中运行标题18 f-ftt目标PARP-1

证据评论:腺苷脱氨酶缺乏2
六篇论文被确定为纳尔木单抗单一疗法的综述。两项研究是回顾性队列研究(Andriessen等,2023年和Li等2023),但被视为回顾性病例序列,以进行分析,作为相关数据,以个人患者为基础。同类的总样本量为29例和30例患者,每项研究的9例患者均接受Adalimumab治疗,并包括在本综述中。其余四项研究是回顾性病例系列(Cooray等,2021,Deuitch等2022,Melo等,2023和Nihira等人2021),其中并非所有患者都处于范围(即一些患者接受了其他TNF抑制剂的治疗),相关数据主要用个别患者进行描述。包括样本量范围为三到十四。目前尚不清楚Andriessen等人(2023),Li等人(2023年和Deuitch等人(2022)中是否存在重叠的患者,因为Andriessen等人(2023)(2023)和Li et al(2023)的一些患者(2023)中属于健康群体的国家研究所。


聚腺苷二磷酸的动力学和静态分析
聚腺苷二磷酸核糖聚合酶 (PARP) 蛋白家族参与多种功能,最显著的是 DNA 损伤反应。癌症易受 DNA 损伤的影响,这导致了几种 PARP 抑制剂 (PARPi) 的开发。这类药物已被证明对卵巢癌、乳腺癌和前列腺癌有治疗效果,但反应各不相同。因此,诊所需要选择可能从这些靶向疗法中受益的患者。体内 18 F-氟邻苯二甲酸酯摄取成像已被证明与组织中的 PARP-1 表达相对应。本研究表征了 18 F-氟邻苯二甲酸酯的药代动力学,并测试了动力学和静态模型,以指导未来研究中评估 18 F-氟邻苯二甲酸酯作为 PARPi 治疗反应生物标志物的指标选择。方法:14 名前瞻性入选的卵巢癌患者注射 18 F-氟苯那敏,注射后进行 60 分钟动态成像,随后进行最多 2 次全身扫描,测量静脉血活性和代谢物。从动态图像和全身扫描中提取 SUV 最大值和 SUV 峰值。评估动力学参数估计值和 SUV 与组织 PARP-1 免疫荧光的相关性(n=7)。群体动力学参数的模拟可以估计测量偏差和参数估计的精度。结果:18 F-氟苯那敏血液清除率各不相同,但不同患者的标记代谢物谱相似,支持使用群体母体分数曲线。可逆性2组织室模型和Logan参考组织分布体积(DVR)在PET采集的第一个小时内的总分布体积与免疫荧光检测的肿瘤PARP-1表达相关(分别为r=0.76和0.83;P=0.05)。DVR偏差和精度估计分别为6.4%和29.1%。从中点为57.5、110±3和199±4分钟的图像获得的SUV max和SUV peak与PARP-1表达高度相关(平均值±SD,r=0.79;P=0.05)。结论:注射后55-60分钟及以后的肿瘤SUV max和SUV peak以及至少60分钟的DVR似乎是PARP-1结合的可靠非侵入性测量方法。 18 F-氟苯那敏在卵巢癌中的吸收最好用可逆结合模型来描述。然而,示踪剂吸收的药代动力学模式有些变化,尤其是在后期。
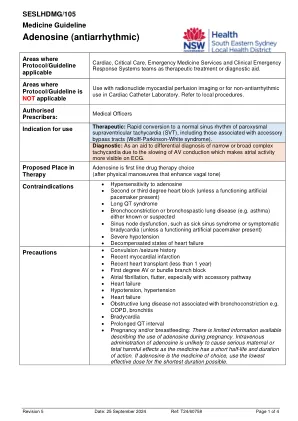
药物指南 - 腺苷(抗心律失常药)
注意事项 • 抽搐/癫痫病史 • 近期心肌梗塞 • 近期心脏移植(不到 1 年) • 一度 AV 或束支传导阻滞 • 心房颤动、扑动,尤其是伴有旁路 • 心力衰竭 • 低血压、高血压 • 心力衰竭 • 与支气管收缩不相关的阻塞性肺病,如慢性阻塞性肺病、支气管炎 • 心动过缓 • QT 间期延长 • 怀孕和/或哺乳:关于怀孕期间使用腺苷的信息有限。由于腺苷的半衰期和作用持续时间较短,因此静脉注射腺苷不太可能对孕妇或胎儿造成严重的有害影响。如果选择腺苷作为药物,请尽可能使用最低有效剂量并缩短用药时间。

用腺苷受体阻滞剂解锁抗肿瘤免疫
摘要肿瘤通过创建抑制抗肿瘤免疫力的肿瘤微环境(TME)而生存。TME通过限制抗原表现,抑制淋巴细胞和天然杀伤(NK)细胞激活并促进T细胞衰竭来抑制免疫系统。抗PD-1和抗CTLA(例如抗PD-1)的检查点抑制剂是免疫刺激性抗体,它们的封锁扩展了一些但不是所有癌症患者的存活率。细胞外腺苷三磷酸(ATP)在发炎的肿瘤中很丰富,其代谢产物腺苷(ADO)是由腺苷A2A受体(A2AR)和腺苷A2B受体(A2BR)介导的免疫抑制作用的驱动器,并且在肿瘤中发现了在肿瘤中的lumor-lymphassiped lympys-symphassiped lympssed Lympsspatied lymphoidsyeloideciped lymphoid和myelyeloideroid。本综述将集中于TME中的关键检查点抑制剂样的免疫抑制剂,以及如何减少腺苷产生或阻断A2AR和A2BR增强抗肿瘤免疫力。

抑制腺苷A2A受体的抗体拮抗剂
森林生态系统储存大量碳,可以是大气二氧化碳的重要来源或下沉,这有助于全球变暖。了解不同森林的碳存储潜力及其对管理和干扰事件的反应是制定政策和场景以部分抵消绿色房屋气体排放的基础。在不同模型中,活树碳积累的投影的处理方式不同,结果不一致。我们开发了增长和收益模型,以从7,523个国家森林库存图中预测沿海太平洋地区(美国加利福尼亚,俄勒冈州和华盛顿)的所有植被类型中的独立型活树碳密度作为展台年龄的函数。我们将场地的生产率和库存性纳入了章节式方程式,并测试了强化管理的私人森林的行为与管理较低的公共森林的行为不同。我们发现,最好的模型将库存性纳入了方程式术语控制架的承载能力中,以及以方程式术语来控制曲线生长速率和形状的现场生产力。RMSE的不同植被类型的范围为10至137 mg c/ha。对生产性的道格拉斯 - 费尔/西部铁杉区的标准工业旋转长度(约50年),所有者没有显着影响,这表明库存性和生产力的差异捕获了归因于管理强度的许多变化。我们的模型表明,将这些强度管理的土地上的旋转长度从35年增加到70年,将导致储存在景观上的活树碳的2.35倍。这些发现与某些研究预计具有相同植被类型的碳密度较高的研究不一致,并且尚未发现(每年)旋转较长的收益率(每年)增加。我们怀疑差异主要是由于用完全储存的,不受干扰的单物种开发的屈服曲线,“正常”的立场,而没有考虑到不符合这些假设的大量森林。此处开发的碳积聚曲线可以直接以生长和收益样式的投影模式应用,并用于验证生态生理学,同类或单树样式的预测