XiaoMi-AI文件搜索系统
World File Search SystemmRNAs

超级金属:一种生成的AI框架,用于蛋白质中的快速,精确的金属离子位置预测
最初的计算方法用于mRNA定位是单个标签分类任务,其中每个mRNA被预测仅定位为一个特定的隔室。rnatracker采用了深层复发的神经网络来预测mRNA定位[6]。iloc-mRNA,利用支持载体机(SVM)来预测在同性恋中的mRNA定位,[7]。sublocep通过集中在特定的细胞室,同时保留在单标签分类框架内[8],进一步完善了预测。但是,它们本质上受到这样的假设,即每个mRNA仅定位到一个与生物学现实不符的隔间。许多mRNA众所周知,可以定位在多个隔间中,从而在细胞内履行各种作用[9,8]。

寻找乳腺癌中的自噬生物标志物
自噬会导致耐药性和药物诱导的癌细胞毒性。针对自噬过程可以大大改善化疗效果。由于在临床环境中难以可靠地测量自噬水平,因此特定抑制剂或激活剂的发现受到了阻碍。我们通过将具有不同 ER/PR/Her2 受体状态的乳腺癌细胞系暴露于已知但不同的自噬诱导剂(每种诱导剂都有独特的分子靶点,即他莫昔芬、曲妥珠单抗、硼替佐米或雷帕霉素)来研究药物诱导的自噬。在自噬通量最早出现时提取的总 RNA 的差异基因表达分析显示了细胞和药物特异性变化。我们分析了差异表达基因列表,以找到一个共同的、与细胞和药物无关的自噬特征。所有药物均显著调节了 12 种 mRNA,其中 11 种通过 Q-RT-PCR 进行了正交验证( Klhl24 、 Hbp1 、 Crebrf 、 Ypel2 、 Fbxo32 、 Gdf15 、 Cdc25a 、 Ddit4 、 Psat1 、 Cd22 、 Ypel3 )。与药物无关的 mRNA 特征同样由线粒体靶向药物 MitoQ 诱导。对 KM-plotter 癌症数据库进行的计算机模拟分析表明,这些 mRNA 的水平在人类样本中是可检测到的,并且与乳腺癌预后结果相关,包括所有患者的无复发生存期 (RSF)、所有患者的总生存期 (OS) 和 ER + 患者的无复发生存期 (RSF ER +)。 Klhl24 、 Hbp1 、 Crebrf 、 Ypel2 、 CD22 和 Ypel3 水平高与更好的结果相关,而 Gdf15 、 Cdc25a 、 Ddit4 和 Psat1 水平低与乳腺癌患者预后较好相关。该基因特征揭示了候选自噬生物标志物,可在临床前和临床研究中进行测试,以监测自噬过程。

microRNA:基因表达的关键调节剂及其治疗潜力。
治疗上的miRNA提出了挑战和机遇。因为它们在全球范围内调节基因表达会改变miRNA活性,这可能会恢复疾病状态的正常细胞功能。但是,一个miRNA可以针对数百个mRNA的miRNA网络的复杂性意味着治疗策略必须高度具体,以避免意外后果。一种方法是开发合成miRNA模拟物或抑制剂。miRNA类似物的设计是为了增加癌症等疾病中表达的miRNA的表达,而miRNA抑制剂可用于阻断过表达的miRNA的活性。研究这些基于miRNA的疗法的临床试验仍处于早期阶段,但作为治疗靶标的miRNA的潜力仍然很重要。研究这些基于miRNA的疗法的临床试验仍处于早期阶段,但作为治疗靶标的miRNA的潜力仍然很重要。


组蛋白/DNA修饰与RNA N
n 6-甲基腺苷(M 6 A)是真核信使RNA(mRNA)中最普遍的内部RNA修饰,在转录和转录后水平调节基因表达。M 6 A与其他良好研究的表观遗传修饰(包括组蛋白修饰和DNA修饰)之间的复杂相互作用已被广泛报道。RNA M 6之间的串扰修饰和组蛋白/DNA修饰在建立基因表达的精确和特定微调的染色质状态和无疑对生理和病理过程都产生深远影响。在这篇综述中,我们讨论了RNA M 6的修改与组蛋白/DNA修饰之间的串扰,强调了它们的复杂通信和基于的机制,以使M 6 A基于M 6的生物学相关性具有全面的看法。
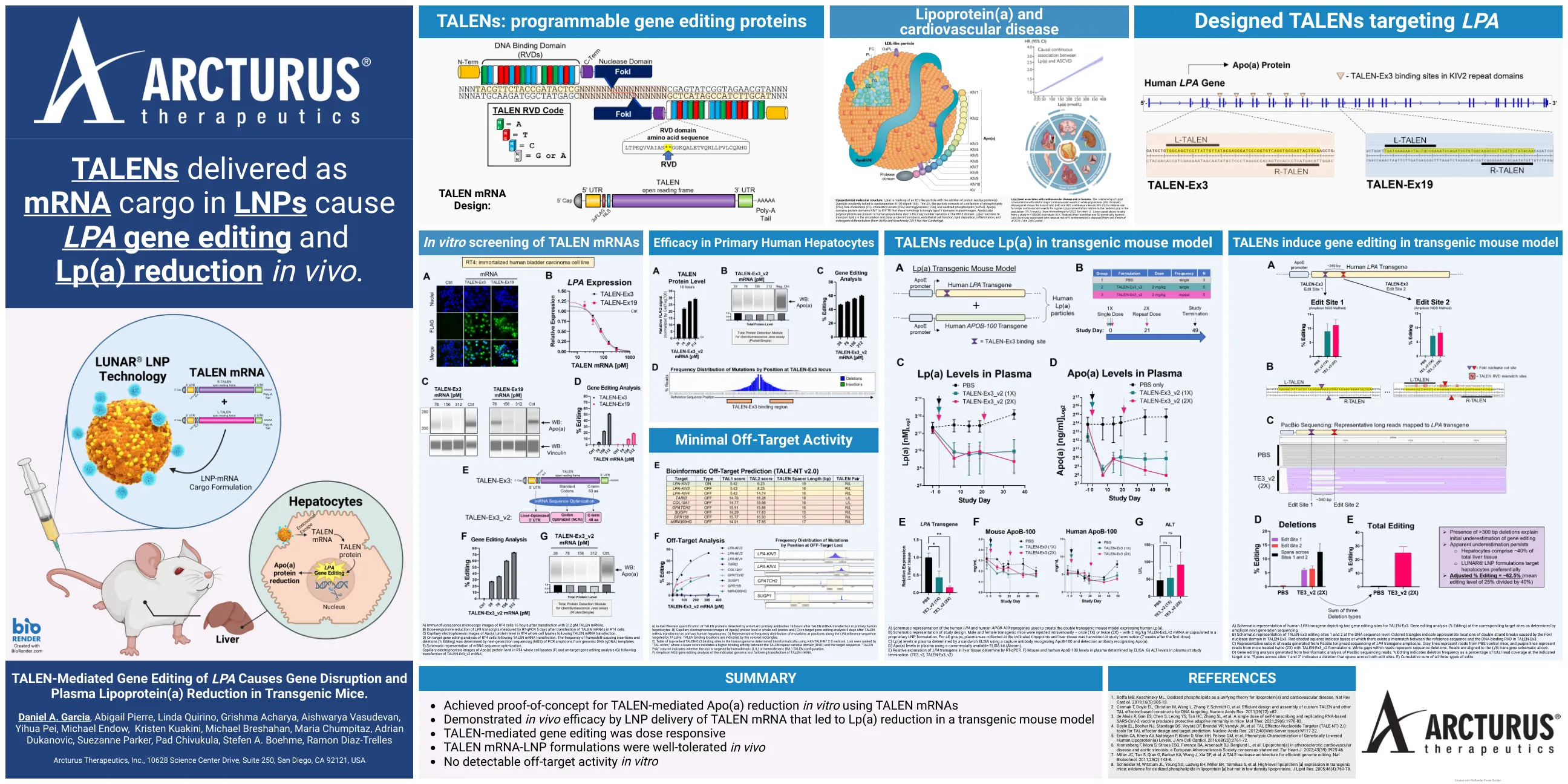
LPA 海报 ASGCT 2023
A) RT4 细胞转染 312 pM TALEN mRNA 16 小时后的免疫荧光显微镜图像。B) RT4 细胞转染 TALEN mRNA 5 天后,通过 RT-qPCR 测量 LPA 转录本的剂量反应性减少。C) TALEN mRNA 转染后 RT4 全细胞裂解物中 Apo(a) 蛋白水平的毛细管电泳图像。D) TALEN mRNA 转染后 RT4 细胞的靶向基因编辑分析。通过基因组 DNA (gDNA) 模板的 PCR 扩增子的下一代测序 (NGS) 确定导致移码的插入和缺失的频率 (编辑百分比)。E) mRNA 序列优化的示意图。转染 TALEN-Ex3_v2 mRNA 后 RT4 全细胞裂解物中 Apo(a) 蛋白水平的毛细管电泳图像 (F) 和靶向基因编辑分析 (G)。

分析方法-RSC发布
在所有生物标志物中,科学家们都越来越关注小小的miRNA(大约长度为22个核苷酸)非编码调节性RNA,可能会通过与mRNA相互作用来影响几乎所有人类疾病的发展和进展。15,16实际上,任何单个miRNA都可以通过广泛的miRNA - mRNA相互作用网络靶向并抑制数百个mRNA,因此会影响广泛的细胞和生物学过程。近年来,miRNA作为一种新型的生物标志物表现出了巨大的希望,用于检测各种疾病,包括癌症,17个疾病中的17种疾病,其中17种在炎性慢性疾病18等。19然而,由于其固有特征引起的某些技术困难,例如

非蛋白质编码的RNA垃圾或宝藏?
图2核糖开关机制,功能和保护。(a)核糖开关是高度结构化的RNA基序,这些RNA基序嵌入了许多细菌mRNA的5'非翻译区域中,在那里它们可以在共同转文时增强或抑制基因表达,以结合小分子或元素离子离子配体。这样的机制涉及RNA聚合酶(RNAP)对转录产量的调节,而其他机制则更直接地改变了mRNA转化为蛋白质的可能性。(b)上游适体区域结合配体,渲染形成结合口袋(黄色框)的核心段以及侧翼建筑片段(蓝色框),高度保守。[112,113]相比,下游表达平台显示出较少的保护,最可能是因为它在功能上与许多对特定细菌具有特殊性的蛋白质效应子相互作用。使用biorender.com创建。

核酸药物研发与靶向治疗
胰腺癌 (PC) 是最致命的癌症之一,5 年生存率接近 10%。由于 PC 具有高度异质性、促纤维化肿瘤微环境和低效药物渗透性,目前推荐用于治疗 PC 的化疗策略临床获益有限。基于核酸的靶向疗法已成为药物发现和靶向治疗领域的强大竞争对手。大量证据表明,基于抗体或适体的策略在很大程度上促进了药物在肿瘤中的积累增加,同时降低了系统性细胞毒性。本综述介绍了反义寡核苷酸 (ASO)、小干扰 RNA (siRNA)、微小 RNA (miRNA)、信使 RNA (mRNA) 和适体-药物偶联物 (ApDC) 在 PC 治疗中的进展,揭示了 PC 治疗的光明应用和发展方向。
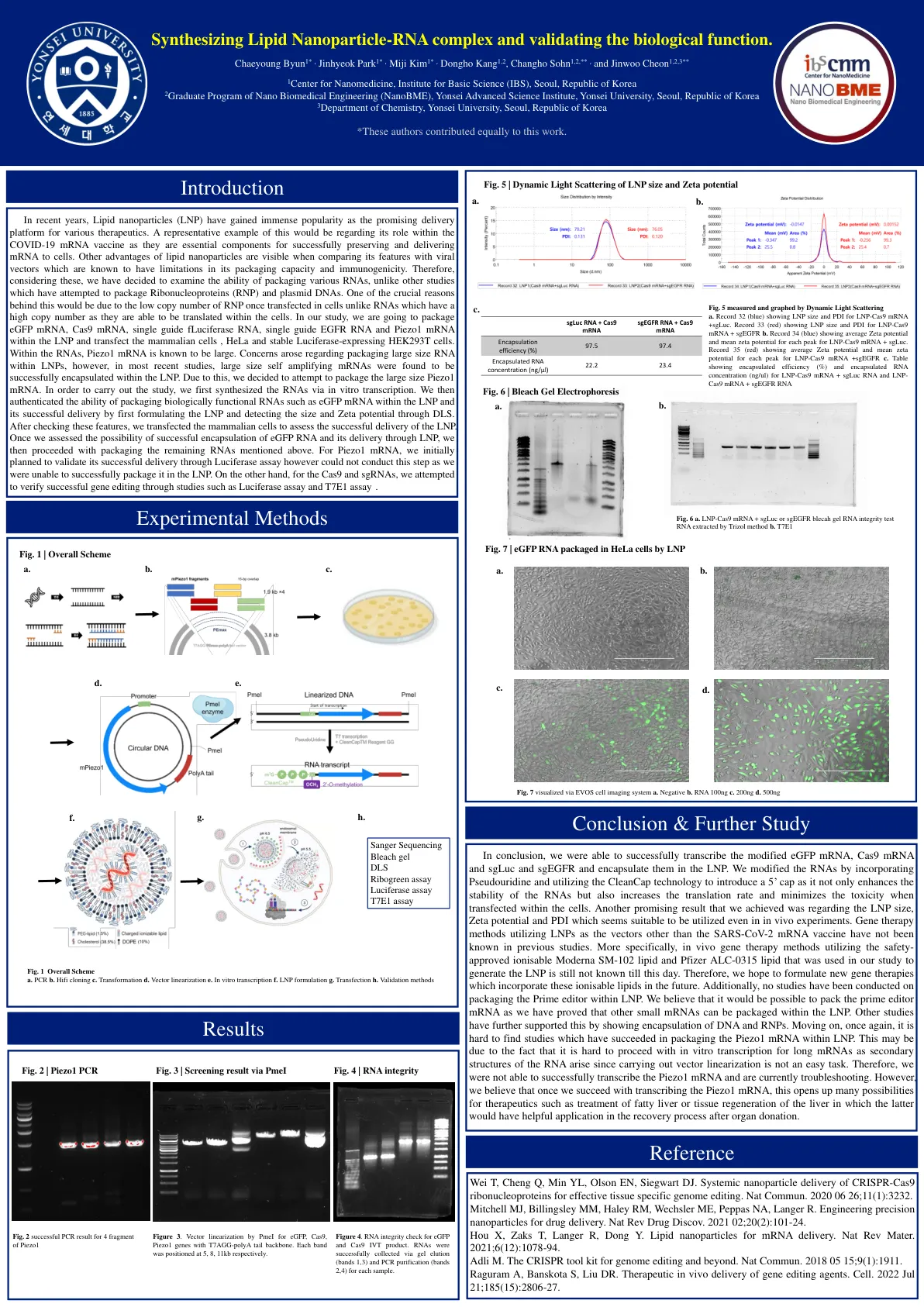
合成脂质纳米粒子-RNA复合物并验证其生物学功能。
总之,我们能够成功转录修饰的 eGFP mRNA、Cas9 mRNA 和 sgLuc 和 sgEGFR 并将它们封装在 LNP 中。我们通过加入假尿苷并利用 CleanCap 技术引入 5' 帽来修饰 RNA,因为它不仅可以增强 RNA 的稳定性,还可以提高翻译率并最大限度地降低在细胞内转染时的毒性。我们取得的另一个有希望的结果是关于 LNP 大小、Zeta 电位和 PDI,这似乎适合在体内实验中使用。以前的研究中还没有发现利用 LNP 作为载体(除 SARS-CoV-2 mRNA 疫苗外)的基因治疗方法。更具体地说,利用安全批准的可电离 Moderna SM-102 脂质和辉瑞 ALC-0315 脂质(我们在研究中用于生成 LNP)的体内基因治疗方法至今仍不得而知。因此,我们希望将来能开发出结合这些可电离脂质的新型基因疗法。此外,目前还没有关于将 Prime editor 包装在 LNP 中的研究。我们相信可以包装 prime editor mRNA,因为我们已经证明其他小 mRNA 也可以包装在 LNP 中。其他研究通过展示 DNA 和 RNP 的封装进一步支持了这一点。再说一遍,很难找到成功将 Piezo1 mRNA 包装在 LNP 中的研究。这可能是因为长 mRNA 的体外转录很难进行,因为 RNA 的二级结构会出现,因为进行载体线性化不是一件容易的事。因此,我们无法成功转录 Piezo1 mRNA,目前正在排除故障。然而,我们相信,一旦我们成功转录 Piezo1 mRNA,这将为治疗开辟许多可能性,例如治疗脂肪肝或肝脏组织再生,后者将有助于器官捐赠后的恢复过程。