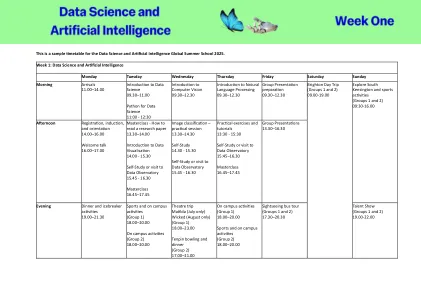
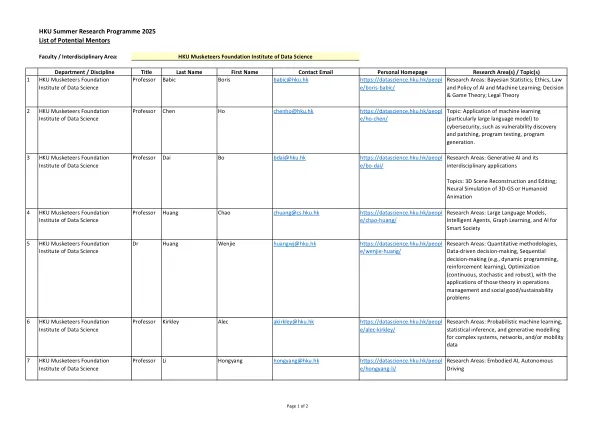
机构名称:
¥ 1.0
基于单倍型的摘要统计数据 - 例如IHS(Voight等人2006),NSL(Ferrer-Admetlla等人 2014),XP-EHH(Sabeti等人。 2007)和XP-NSL(Szpiech等人 2021) - 在进化基因组学研究中司空见惯,以确定种群中的最新和持续的阳性选择(例如, Colonna等。 2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021)。 当适应性等位基因扫描一个人群时,它留下了长期高频单倍型和等位基因附近遗传多样性低的特征模式。 这些统计数据旨在通过总结单倍型纯合性的衰减来捕获这些信号,这是一个距离被推定的区域(IHS和NSL)或两个种群(XP-EHH和XP-NSL)之间的距离。 这些基于单倍型的统计数据非常有力地检测最近的阳性选择(Colonna等人 2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2006),NSL(Ferrer-Admetlla等人2014),XP-EHH(Sabeti等人。 2007)和XP-NSL(Szpiech等人 2021) - 在进化基因组学研究中司空见惯,以确定种群中的最新和持续的阳性选择(例如, Colonna等。 2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021)。 当适应性等位基因扫描一个人群时,它留下了长期高频单倍型和等位基因附近遗传多样性低的特征模式。 这些统计数据旨在通过总结单倍型纯合性的衰减来捕获这些信号,这是一个距离被推定的区域(IHS和NSL)或两个种群(XP-EHH和XP-NSL)之间的距离。 这些基于单倍型的统计数据非常有力地检测最近的阳性选择(Colonna等人 2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2014),XP-EHH(Sabeti等人。2007)和XP-NSL(Szpiech等人2021) - 在进化基因组学研究中司空见惯,以确定种群中的最新和持续的阳性选择(例如,Colonna等。2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021)。 当适应性等位基因扫描一个人群时,它留下了长期高频单倍型和等位基因附近遗传多样性低的特征模式。 这些统计数据旨在通过总结单倍型纯合性的衰减来捕获这些信号,这是一个距离被推定的区域(IHS和NSL)或两个种群(XP-EHH和XP-NSL)之间的距离。 这些基于单倍型的统计数据非常有力地检测最近的阳性选择(Colonna等人 2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2014,Zoledziewska等。2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021)。 当适应性等位基因扫描一个人群时,它留下了长期高频单倍型和等位基因附近遗传多样性低的特征模式。 这些统计数据旨在通过总结单倍型纯合性的衰减来捕获这些信号,这是一个距离被推定的区域(IHS和NSL)或两个种群(XP-EHH和XP-NSL)之间的距离。 这些基于单倍型的统计数据非常有力地检测最近的阳性选择(Colonna等人 2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2015,Ne´de´lec等。2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021)。 当适应性等位基因扫描一个人群时,它留下了长期高频单倍型和等位基因附近遗传多样性低的特征模式。 这些统计数据旨在通过总结单倍型纯合性的衰减来捕获这些信号,这是一个距离被推定的区域(IHS和NSL)或两个种群(XP-EHH和XP-NSL)之间的距离。 这些基于单倍型的统计数据非常有力地检测最近的阳性选择(Colonna等人 2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2016,Crawford等。2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021)。 当适应性等位基因扫描一个人群时,它留下了长期高频单倍型和等位基因附近遗传多样性低的特征模式。 这些统计数据旨在通过总结单倍型纯合性的衰减来捕获这些信号,这是一个距离被推定的区域(IHS和NSL)或两个种群(XP-EHH和XP-NSL)之间的距离。 这些基于单倍型的统计数据非常有力地检测最近的阳性选择(Colonna等人 2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2017,Meier等。2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021)。 当适应性等位基因扫描一个人群时,它留下了长期高频单倍型和等位基因附近遗传多样性低的特征模式。 这些统计数据旨在通过总结单倍型纯合性的衰减来捕获这些信号,这是一个距离被推定的区域(IHS和NSL)或两个种群(XP-EHH和XP-NSL)之间的距离。 这些基于单倍型的统计数据非常有力地检测最近的阳性选择(Colonna等人 2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2018,Lu等。2019,Zhang等。 2020,Salmo´n等。 2021)。 当适应性等位基因扫描一个人群时,它留下了长期高频单倍型和等位基因附近遗传多样性低的特征模式。 这些统计数据旨在通过总结单倍型纯合性的衰减来捕获这些信号,这是一个距离被推定的区域(IHS和NSL)或两个种群(XP-EHH和XP-NSL)之间的距离。 这些基于单倍型的统计数据非常有力地检测最近的阳性选择(Colonna等人 2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2019,Zhang等。2020,Salmo´n等。2021)。当适应性等位基因扫描一个人群时,它留下了长期高频单倍型和等位基因附近遗传多样性低的特征模式。这些统计数据旨在通过总结单倍型纯合性的衰减来捕获这些信号,这是一个距离被推定的区域(IHS和NSL)或两个种群(XP-EHH和XP-NSL)之间的距离。这些基于单倍型的统计数据非常有力地检测最近的阳性选择(Colonna等人2014,Zoledziewska等。 2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2014,Zoledziewska等。2015,Ne´de´lec等。 2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2015,Ne´de´lec等。2016,Crawford等。 2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2016,Crawford等。2017,Meier等。 2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2017,Meier等。2018,Lu等。 2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。 2021)。 此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2018,Lu等。2019,Zhang等。 2020,Salmo´n等。 2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。2019,Zhang等。2020,Salmo´n等。2021),并且两个人群版本甚至可以在很大的参数空间上进行成对的FST扫描(Szpiech等人。2021)。此外,基于单倍型的方法也已证明对背景选择是可靠的(Fagny等人。2014,Schrider 2020)。 然而,这些统计数据中的每一个都认为单倍型相是已知或据估计的。 作为非模型生物的基因组测序数据的产生正在变得常规(Ellegren 2014),有很多很大的机会来研究整个生命之树的最新适应性(例如, Campagna和Toews 2022)。 但是,这些生物/种群通常没有特征良好的人口历史或重组率2014,Schrider 2020)。然而,这些统计数据中的每一个都认为单倍型相是已知或据估计的。作为非模型生物的基因组测序数据的产生正在变得常规(Ellegren 2014),有很多很大的机会来研究整个生命之树的最新适应性(例如,Campagna和Toews 2022)。但是,这些生物/种群通常没有特征良好的人口历史或重组率
selscan 2.0:在未经遗传数据中扫描扫描
主要关键词





相关文件推荐