XiaoMi-AI文件搜索系统
World File Search Systemmutations

基因突变很重要
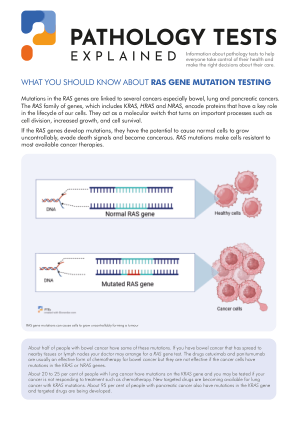
结直肠癌 (CRC) 是全球癌症相关死亡的第三大常见原因,每年有近 100 万人死于该病 (1)。大约一半的转移性 CRC 携带 KRAS(Kirsten 大鼠肉瘤病毒致癌基因同源物)激活突变,导致 GTP 结合活性形式和 GDP 结合非活性形式之间的稳态平衡被破坏。RAS 活性形式的持续存在与上游 RTK 的影响完全脱节,导致主要涉及细胞增殖和迁移过程的几种下游通路过度激活 (2,3)。因此,以 RTK 为靶点的药物(如抗表皮生长因子受体 (EGFR) 单克隆抗体 (moAb))无效。外显子 2 上的密码子 12 和 13 以及外显子 3 上的密码子 61 是最常见的 KRAS 突变位点,而外显子 4 上的密码子 117 和 146 以及其他 RAS 家族成员 HRAS 和 NRAS 上的突变则非常罕见(4-7)。对转移性 CRC 患者中 KRAS 突变的临床影响的理解始于外显子 2 突变被确定为对西妥昔单抗和帕尼单抗等抗 EGFR 单抗反应的负面预测因子(8、9)。然后,对 KRAS 的扩展评估

视紫红质突变的基因治疗
视紫红质基因 RHO 的突变是常染色体显性视网膜色素变性 (adRP) 的很大一部分原因。患者在临床上分为两类:一类是早发性全视网膜光感受器变性,另一类是病情缓慢进展的患者。后一类患者适合接受光感受器定向基因治疗,而前一类患者则适合将光反应蛋白递送至中间神经元或视网膜神经节细胞。RHO adRP 的基因治疗可能针对 DNA 或 RNA 水平的突变基因,而其他疗法则保留光感受器的活力而不解决潜在的突变。在动物模型中,纠正 RHO 基因和替换突变 RNA 显示出良好的前景,而维持可行的光感受器有可能延缓中央视力的丧失,并可能保留光感受器以进行基因定向治疗。

植物中 CRISPR 诱导突变的选择
在本研究中,我们描述了敲除标记基因 MAR1 的建立,用于在组织培养中选择 CRISPR/Cas9 编辑的拟南芥幼苗和番茄外植体。MAR1 编码一种位于线粒体和叶绿体中并参与铁稳态的转运蛋白。它还会随机将氨基糖苷类抗生素转运到这些细胞器中,而该基因的缺陷会导致植物对这些化合物不敏感。在这里,我们展示了由 CRISPR 系统诱导的 MAR1 突变使拟南芥植物和番茄组织具有卡那霉素抗性。MAR1 在多种植物物种中都是单拷贝的,相应的蛋白质形成一个独特的系统发育进化枝,从而可以轻松识别不同植物中的 MAR1 直系同源物。我们证明,在多重方法中,通过由 MAR1 突变介导的 CRISPR/Cas9 诱导的卡那霉素抗性来选择拟南芥幼苗,观察到第二个靶基因突变的频率高于仅因存在转基因而选择的对照群体。这种所谓的共同选择以前从未在植物中发生过。该技术可用于选择经过编辑的植物,如果编辑事件很少发生,这可能特别有用。

人类衰老中的躯体突变
体细胞突变的积累是癌症的驱动力,长期以来一直与衰老有关。由于量化突变负担与非癌组织年龄的限制,尚不清楚体细胞突变对其他衰老表型的影响。DNA测序技术的最新进展允许对衰老组织中的体细胞突变进行大规模定量。这些研究表明,随着年龄的增长,正常组织中突变的逐渐积累,以及主要由癌症相关突变驱动的大量克隆膨胀。然而,很难想象到目前为止确定的与年龄相关的体细胞突变的负担和随机性质如何解释大多数逐渐发展的老化表型。跨物种的研究还发现,寿命较长的物种具有较低的体细胞突变率,尽管这可能是由于对其他表型(例如癌症)作用的选择性压力所致。对较高的体细胞突变负担且没有加速衰老的迹象的最新研究进一步质疑体细胞突变在衰老中的作用。总体而言,除了少数例外,例如癌症,最近的DNA测序研究和遗传突变并不支持这样的观念,即体细胞突变会随着年龄的驱动衰老表型积聚,以及在衰老中的体细胞突变(如果有的话)仍然不清楚。

糖尿病中OneCut1的突变和变体
对应作者:Albert-Einstein-Allee Internechance I Alexander Kleger教授,Albert-Einstein-Allee 23,89081 Ulm,德国,电话: +49-731-500-44728,传真: +49-731-731-500-444612,Alexander.klegleni-umi-ulm.dee; CécileJulier,内分泌学,代谢和糖尿病系,科钦研究所,24 Rue du Faubourg Saint-Jacques,75014法国巴黎,电话:+33.1.44.41.41.22.33#这些作者为这项工作做出了同样的贡献。*这些作者也同样贡献。作者贡献AP,SH,IGC和VS被获取,分析和解释数据,起草和修改工作。AP对项目的人类遗传部分进行了统计,遗传和生物信息学分析。SH对RNA,ATAC-SEQ和质谱法进行了PSC和准备样品的功能研究。IGC对项目进行了并定向生物信息学分析。与糖尿病患者及其家人以及德国队列的测序和基因分型。MB,ZL和GK获得了数据并进行了数据分析。ad,pz,hn,ES,TK,MW,CB,RO,JFD,BK,CDR获得了该项目的数据。MB,RG,MHE和TS修订了手稿。具体来说,MB和AI进行了hESC和初始功能分析的基因编辑。Zl,GK和XZ进行了chip-seq,Zl,GK和MSC进行ATAC-SEQ和ZL,GK,MSC和QL RNA-SEQ生物信息信息分析。RR获得了数据并对工作进行了大量修订。MHO对患者成纤维细胞和IPSC分析进行了重编程。SL解释了数据并修改了工作。JRB生成的记者ESC线路。对WES数据进行了生物信息学分析,并在临床上描述了黎巴嫩患者及其家人的PZ。MSA解释了提供的数据,提供了材料,修改了工作。JK获得并分析了质谱数据。AW awed并提供了RG提供芯片序列数据。 kg,JC解释了遗传学数据,而GN提供了来自分化MEL1 HESC的RNA。 Bob,FO,MN,CJ和AK负责获取和分析数据的起草和修订。 此外,鲍勃(Bob)也指导了有关德国患者队列和解释遗传学数据的研究,FO表达和分析了TFS和ONECUT1变体,MN确定并临床表征了患者1及其大家庭,并解释了人类的遗传和临床数据。 此外,CJ和AK设计了工作,解释了数据并用所有作者的输入起草了手稿。 cj指导项目的遗传部分,并进行了人类遗传分析。 AK指导该项目的功能研究。AW awed并提供了RG提供芯片序列数据。kg,JC解释了遗传学数据,而GN提供了来自分化MEL1 HESC的RNA。Bob,FO,MN,CJ和AK负责获取和分析数据的起草和修订。此外,鲍勃(Bob)也指导了有关德国患者队列和解释遗传学数据的研究,FO表达和分析了TFS和ONECUT1变体,MN确定并临床表征了患者1及其大家庭,并解释了人类的遗传和临床数据。此外,CJ和AK设计了工作,解释了数据并用所有作者的输入起草了手稿。cj指导项目的遗传部分,并进行了人类遗传分析。AK指导该项目的功能研究。

DMD 突变的复杂状况
杜氏肌营养不良症 (DMD) 是一种严重的遗传性疾病,其特征是进行性肌肉退化,伴有呼吸和心脏并发症,是由编码蛋白质肌营养不良蛋白的 DMD 基因突变引起的。各种 DMD 突变会导致不同的表型和疾病严重程度。了解基因型/表型相关性对于优化临床护理至关重要,因为突变特异性疗法和创新治疗方法正在变得可用。疾病修饰基因,影响疾病严重程度和表型表现的反式活性变体,可能会调节对治疗的反应,并成为新的治疗靶点。通过广泛的基因组图谱研究发现更多的疾病修饰基因,有可能为患有 DMD 的个体微调预后评估。本综述提供了对基因型/表型相关性和修饰基因在 DMD 中的影响的见解。


肝细胞倍性和病理突变...
肝细胞癌 (HCC) 是一种原发性肝脏肿瘤,是日本第五大癌症死亡原因。大多数 HCC 见于肝硬化或慢性肝损伤患者,例如病毒感染[乙型肝炎病毒 (HBV)、丙型肝炎病毒 (HCV)]、酒精性损伤、非酒精性脂肪性肝病 (NAFLD) 以及包括原发性胆汁性胆管炎和自身免疫性肝炎在内的自身免疫性疾病。WHO 估计 53% 的 HCC 发生在 HBV 感染患者中,另有 25% 发生在 HCV 感染患者中。相反,在日本,大约 65% 的 HCC 病例是由 HCV 感染引起的,15% 由 HBV 感染引起(1)。日本最近的多机构全国性调查报告称,因饮酒或非酒精性脂肪性肝炎 (NASH) 引起的非病毒性肝硬化的比例有所增加(2)。为了根据肝脏的局部情况预防慢性损伤,可以采取控制病毒感染的措施,以及改变生活方式,包括减少饮酒、健康饮食和体育锻炼,作为二级预防。尽管 HCV 的直接抗病毒 (DAA) 疗法和 HBV 的核苷酸类似物已被广泛使用,但 HCC 仍然是少数几种

在体内抑制tonb盒的tonb盒共识pentapeptide
大肠杆菌不匹配维修系统能够识别DNA中的非分配基础对,显然是通过局部切除和重新合成的,以取代错误的基础(有关审查,请参见参考,请参见参考文献1)。DNA的区域GATC序列是完全腺嘌呤 - 甲基化的似乎是对不匹配修复的难治性(2,3),并且似乎是在复制叉后紧接在复制后立即将新合成的GATC序列的短暂甲基化,从而使修复的重复修复仅可重复进行新的合成,从而将其撤离了新的合成和错误。大肠杆菌不匹配修复系统没有识别和/或维修所有不匹配的效率(6,7)。两个过渡不匹配(G-T和CGA)都很容易予以修复和修复,而六个转移不匹配中的三个不是(6)。这种模式可以部分解释,因为发现在大肠杆菌,mutl,muts和mutu突变体中观察到的突变效应,这些突变体缺乏不匹配修复(参考文献。2-8;有关评论,请参见参考。1)和未指向不匹配修复的大坝突变体(2,6)主要是由于过渡和移码突变的增加(1)。不匹配维修不足的突变体显示移码突变的频率增加,这表明大肠杆菌不匹配修复系统可以识别和修复一个或多个未配对的碱基 - i.e。,移交/野生型型异源杂质。该假设进行了检验。结果表明,具有一个未配对基碱的异源型可以通过大肠杆菌不匹配修复系统识别和修复。