机构名称:
¥ 2.0
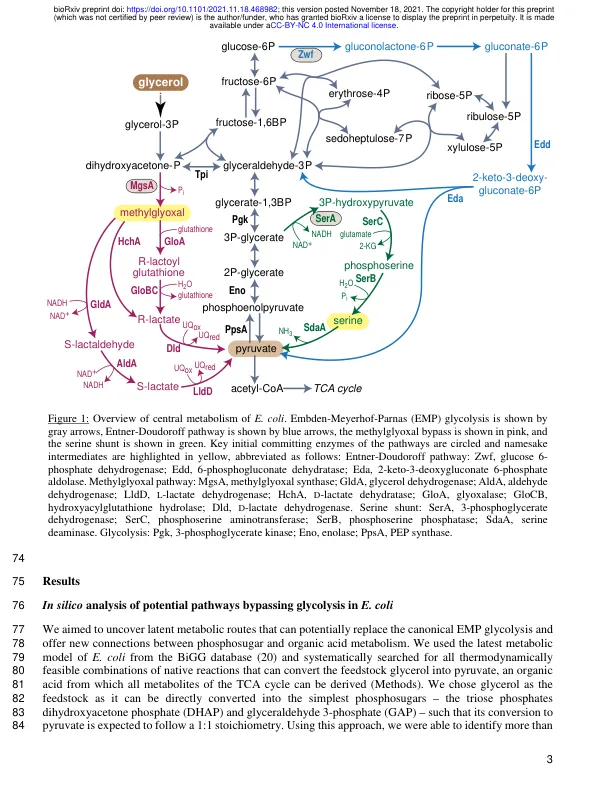
所有活生物体在其中央代谢中都有类似的反应,为所有19个基本构件和降低力量提供了前体。确定糖酵解20的替代代谢途径是否可以在大肠杆菌中运行,我们在硅设计,合理的工程和自适应21实验室进化中互补。首先,我们使用了一个基因组规模模型,并在该生物体的22个代谢网络中鉴定了两种潜在途径,取代了规范的Embden-Meyerhof-Parnas(EMP)糖酵解,将23个转化为有机酸的磷酸化。这些糖酵解路线之一是通过甲基乙二醇(通过丝氨酸生物合成和降解)进行的。然后,我们在大肠杆菌菌株中实施了两种途径25具有缺陷的EMP糖酵解。令人惊讶的是,通过甲基乙二醇的途径立即在26个三氧磷酸异构酶缺失菌株中培养在甘油上。相比之下,在磷酸甘油酸激酶27缺失菌株中,对于实现功能性28甲基甘氨酸途径的过表达是必要的。此外,我们设计了“丝氨酸分流”,该“丝氨酸分流”通过丝氨酸生物合成和降解转换为丙酮酸,绕过烯醇酶缺失。最后,为了探索30种这些替代方案中的哪些替代方法,我们使用烯醇酶缺失菌株进行了自适应实验室31进化研究。证明进化的突变体使用丝氨酸分流。32我们的研究揭示了代谢途径的灵活性重新定位,以建立新的代谢产物链接和重新连接33中央代谢。34
激活大肠杆菌中的无声糖酵解旁路
主要关键词




相关文件推荐