机构名称:
¥ 1.0
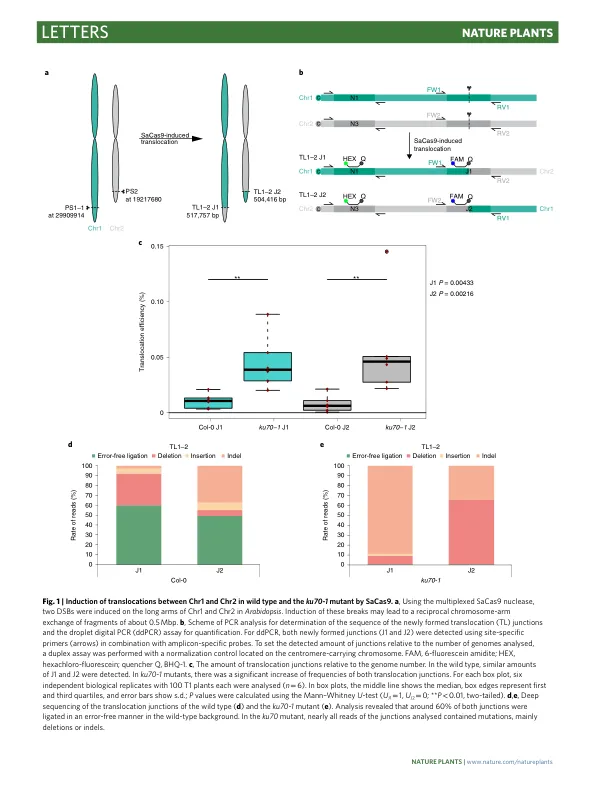
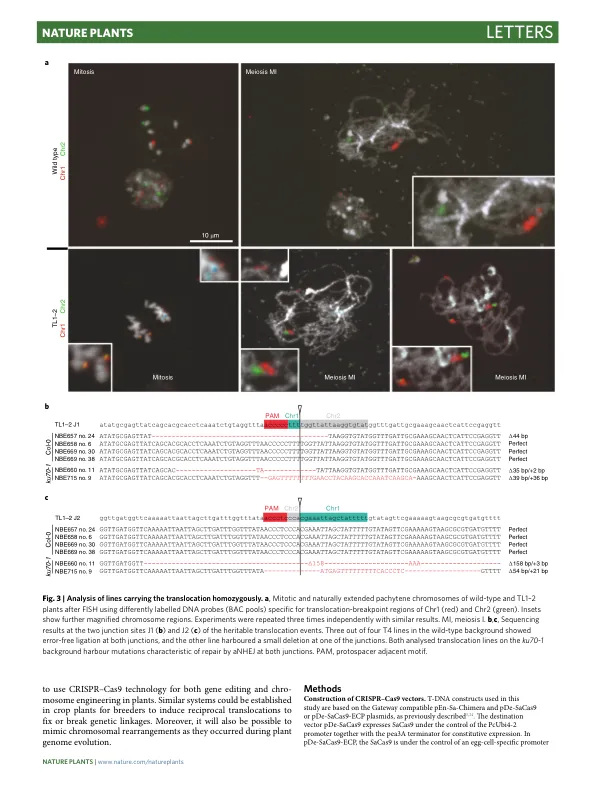
成簇的规律间隔短回文重复序列 (CRISPR) - CRISPR 相关蛋白 (Cas) 技术已应用于植物育种,主要用于改良单个或多个性状的基因 1 – 4 。本文我们表明,这项技术还可用于重组植物染色体。利用来自金黄色葡萄球菌 5 的 Cas9 核酸酶,我们能够在拟南芥中诱导异源染色体之间 Mbp 范围内的相互易位。值得注意的是,在没有经典的非同源末端连接途径的情况下,易位频率大约高出五倍。利用 Cas9 核酸酶的卵细胞特异性表达和连续的批量筛选,我们能够分离可遗传事件并建立易位纯合的品系,单个品系的频率高达 2.5%。通过分子和细胞学分析,我们证实了在拟南芥 1 号和 2 号染色体之间以及 1 号和 5 号染色体之间获得的染色体臂交换是保守的和相互的。诱导染色体易位可以有针对性地模拟基因组进化或染色体修改,固定或打破不同染色体上性状之间的遗传连锁。植物基因组的受控重组有可能改变植物育种。鉴于养活快速增长的人口的挑战以及气候变化对农业的影响,对新作物品种的需求日益增加。随着传统育种已达到极限,使用基因组编辑工具对作物进行理想性状改造正成为主要关注点 6 。应用 CRISPR-Cas 系统定向诱导位点特异性双链断裂 (DSB) 使得基因编辑既可用于植物基础研究,也可用于农业性状的产生和改良 7 。在包括植物在内的多细胞真核生物中,DSB 的修复主要由两种途径介导,非同源末端连接 (NHEJ) 和同源重组 8 。通过易错的 NHEJ 进行的修复通常与断裂位点处的序列信息丢失有关,而同源重组主要导致无错修复 9 。在植物中,NHEJ 是体细胞组织中普遍的修复途径。NHEJ 可进一步细分为经典 NHEJ (cNHEJ) 和替代 NHEJ (aNHEJ) 途径 10 。在 cNHEJ 的情况下,断端直接重新连接,有时会导致断裂位点处的小插入或缺失 (indel)。aNHEJ 利用靠近断裂位点的微同源性并依赖于聚合酶 theta,导致与插入部分相关的微同源性之间的序列信息缺失 11,12 。一次诱导多个 DSB 可以通过 NHEJ 将不相关的断裂末端连接起来,从而导致基因组中复杂的重排。
CRISPR–Cas9 介导的拟南芥可遗传染色体易位的诱导
主要关键词



相关文件推荐