
机构名称:
¥ 1.0
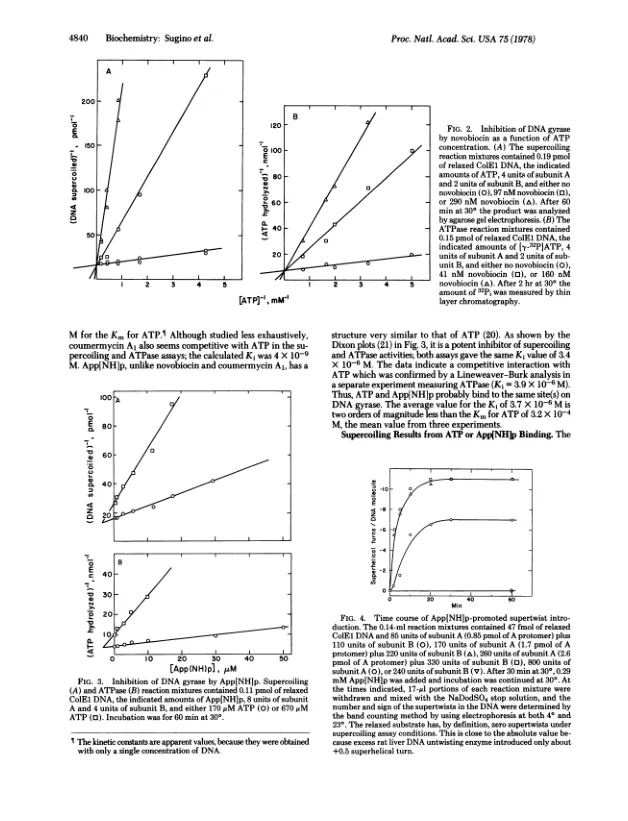
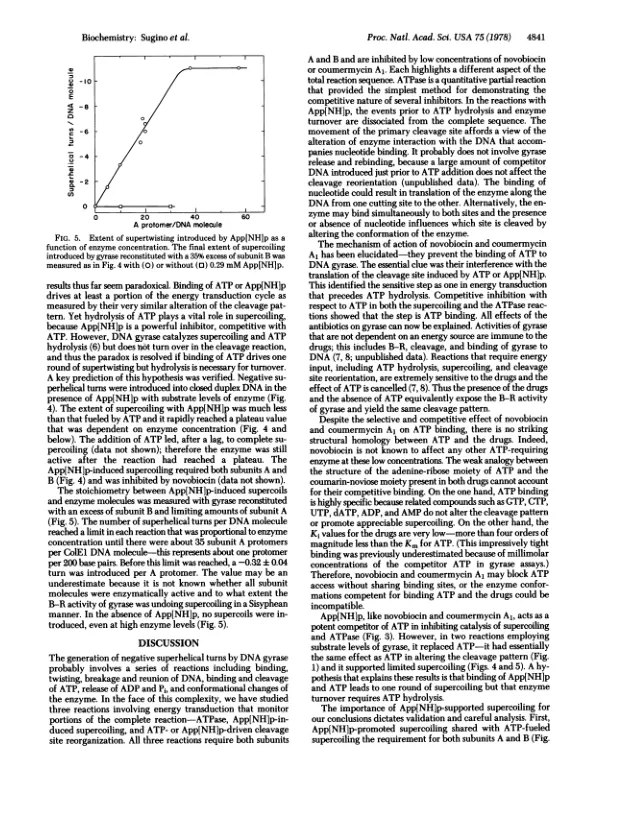
抽象的大肠杆菌DNA速酶催化封闭的双链DNA的否定性超涂层,以ATP为代价。酶的酶的另外活性阐明了超涂层反应的能量偶联成分是ATP至ADP和ADP和PI的DNA依赖性水解,以及ATP通过gyrase裂解反应的DNA位点特异性的ATP改变。这两种DNA链的这种裂解是由稳定的Gy- Rase-DNA复合物的十二烷基硫酸钠处理的,该配合物被抑制剂氧甲酸捕获。ATP或不可水解的类似物,腺基-5'-二氨基磷酸酯(APP [NHLP),都会在Colel DNA上移动主要的裂解位点。这种切割重排的Novobiocin和Coumermycin al的预防将抗生素的作用位点放置在ATP水解之前的一个反应步骤中。步骤阻塞是ATP的结合,因为香豆素和Novobiocin在ATPase和SuperCoiling分析中与ATP竞争相互作用。 K;对于ATP而言,值比KM少四个数量级以上。这种简单的机制解释了药物对DNA回旋酶的所有影响。使用APP [NHP [NHP的另一种有效的反应竞争抑制剂催化YGYRASE的竞争抑制剂,表明将DNA驱动到更高的能量超胶结形式不需要高能键的裂解。 与Gyrase,App的底物水平(NHLP诱导与酶量成正比的超串联; a -0.3超螺旋转弯是根据Gyrase Frotomer A引入的。 我们假设ATP和APP [NH] P是回旋酶的构象变化的变构效应器,导致一轮超涂层。使用APP [NHP [NHP的另一种有效的反应竞争抑制剂催化YGYRASE的竞争抑制剂,表明将DNA驱动到更高的能量超胶结形式不需要高能键的裂解。与Gyrase,App的底物水平(NHLP诱导与酶量成正比的超串联; a -0.3超螺旋转弯是根据Gyrase Frotomer A引入的。我们假设ATP和APP [NH] P是回旋酶的构象变化的变构效应器,导致一轮超涂层。通过ATP水解的核苷酸解离,将回旋酶返回其原始构型,从而允许酶转移。伴随核苷酸亲和力改变的这种环状构象变化似乎也是其他多种操作中能量转导的共同特征,包括肌肉收缩,蛋白质合成和氧化磷酸化。
DNA回旋酶中的能量耦合和作用机理
主要关键词



相关文件推荐





























