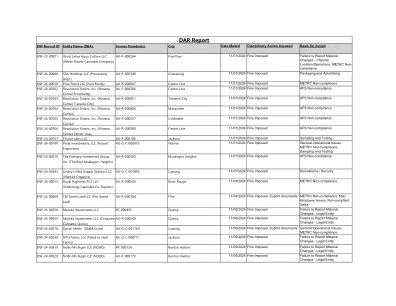
机构名称:
¥ 1.0
含有假定的 G-四链体形成序列的寡核苷酸(PQS;G ≥ 3 N x G ≥ 3 N x G ≥ 3 N x G ≥ 3)在阳离子存在下的生理缓冲条件下(Bochman 等人,2012 年)。由于其高热力学稳定性,组装的 G4 需要通过酶促分解。已经开发出体外用于监测 G4 形成的方法(Balasubramanian 等人,2011 年;Bryan 和 Baumann,2011 年)。使用这些方法已经证明了分解 G4 的酶活性。这些酶包括具有 G4 结合和解旋活性的 DNA 解旋酶,例如 BLM、WRN、PIF1、FANCJ、XPD、DNA2 和 RTEL1(Bochman 等人,2012 年;Maizels,2015 年)。使用计算机分析或荧光成像、免疫沉淀或 pull-down 实验来预测体内 G4 的形成,使用有价值的工具 - 例如特异性识别 G4 的免疫球蛋白和单链可变片段 (scFv) (Henderson 等人,2013)、G4 结合化合物 (Mendoza 等人,2016) 或 G4 结合蛋白 (Maizels,2015)。使用这些工具,可以通过免疫沉淀或针对纯化的基因组 DNA 或染色质的 pull-down 来识别 G4 位点,并且这些位点中的很大一部分重现了 PQS (Chambers 等人,2015;Hänsel-Hertsch 等人,2016;Lam 等人,2013;Muller 等人,2010)。 PQS 在基因的调控区(例如启动子、内含子或非翻译区 [UTR])中过度表达,包括致癌基因、重复区(例如端粒和 rDNA)和复制起点 (Maizels & Gray, 2013 )。使用抗体在人类细胞中进行的全基因组 G4 映射揭示了 G4 存在于基因调控区和端粒中 (Hänsel-Hertsch et al., 2016 ; Liu et al., 2016 )。许多 G4 被映射在转录起始位点周围,G4 形成的频率与相应基因的转录水平呈正相关 (Spiegel et al., 2021 ; Zheng et al., 2020 )。使用抗体对 G4-DNA 进行荧光标记,显示细胞核或染色体上存在颗粒状信号;一些信号位于端粒或着丝粒上 (Biffi et al., 2013; Henderson et al., 2013)。使用荧光标记化合物对 G4- DNA 进行可视化,可显示位于核仁中的较大信号,以及位于细胞核中的一些较小信号 (Rodriguez et al., 2012),或整个细胞核中均匀分布的信号 (Shivalingam et al., 2015)。然而,人们对使用体内成像获得的许多未表征信号的亚细胞或基因组位置了解甚少。越来越多的证据表明,在基因体内或周围形成的 G4 通过促进或抑制转录来调节基因活性 (Bochman et al., 2012; Mendoza et al., 2016)。尽管具有这些生物学含义,但 G4 在空间上阻碍了 DNA 复制和转录 (Bochman et al., 2012; Maizels, 2015)。这些生物事件的拖延会增加基因毒性损害的风险;G4 结构清除不足可能
DHX36 通过解开 G‐ 来维持基因组完整性...
主要关键词




相关文件推荐