机构名称:
¥ 3.0
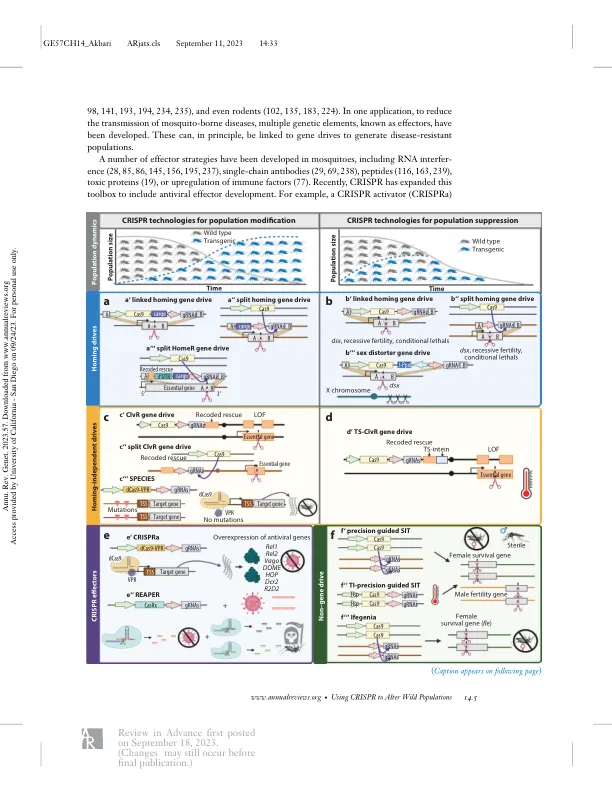
遗传害虫管理策略在 20 世纪初被提出,并于 20 世纪中期开始实施,其中昆虫不育技术 (SIT) 是其中的佼佼者 (130、131、202)。在 SIT 中,不育雄性被释放出来与野生雌性交配,随着时间推移,这种技术频繁大规模释放,可以抑制甚至消灭种群。该领域的早期工作依赖于辐射来产生不育突变 (17、131、207)。大规模实施该技术取得了巨大成功,彻底消灭了北美大部分地区的新大陆螺旋蝇 (131),并抑制了其他一些物种 (83、179)。然而,遗传和其他技术挑战阻碍了抑制某些物种的尝试取得成功。在开展这项工作的同时,人们探索了许多其他控制方法,这些方法基于转基因时代之前对害虫遗传学的操作(例如易位和倒位),但总体上并没有取得很大的成功(100)。人们开始思考用于种群管理的遗传技术,特别是那些旨在自我维持的技术,这种思考始于 50 多年前(64, 201),其灵感来自于生命各个领域中越来越多的自然发生的自私遗传元素 [以下称为基因驱动 (120)] 的行为。许多这样的基因驱动是在遗传学领域早期发现的,通常是由于意外的突变率、性别比例偏差或特定基因型的死亡率而偶然发现的。这些驱动有利于它们的传播,而牺牲了基因组中的其他基因。这种行为可能导致这些驱动相对于相应的染色体对应物扩散,即使它们的存在会给携带者带来适应度成本(即降低整个种群的适应度)(78、95、104、178、226)。自然产生的基因驱动在形式和机制上千差万别,包括性别比例扭曲元件、减数分裂驱动元件和毒素-解毒剂系统(3、66、67、104、117、148)、转座元件(157、178、188)、可遗传微生物(62、80、225)和归巢内切酶(37、38)。这些自然基因驱动的潜在机制启发了合成基因驱动系统的创建(120)。
利用 CRISPR 操控野生种群的命运
主要关键词




相关文件推荐